Cold-water corals and sponges
Writing team: Erik Cordes (coordinating author), Narissa Bax, Jorge Cortes, Sebastien Hennige, Kerry Sink, Joshua Tuhumwire (co-lead member) and Juying Wang (lead member).
Key points
- Exploration of the oceans has revealed the nearly ubiquitous presence of cold-water coral gardens, reefs and sponge grounds on hard substrata throughout the world.
- There is increasing evidence of profound changes in the physical and chemical environment of the deep ocean, threatening cold-water coral and sponge (CWCS) habitats.
- Offshore industrial development continues to pose threats to CWCS habitats globally, with emerging threats from deep-sea mining and marine carbon dioxide (CO2) removal.
- Additional CWCS are being discovered and protected, and are future targets for protections and the development of restoration strategies.
- Ecosystem services-based management is a new philosophy emerging from work in these areas, with a particular focus on the ability of CWCS habitats to sequester carbon on a large scale.
1. Introduction
The present subchapter will focus on CWCS, the habitats they form and the services they provide in the deep sea (at depths of below 200 m). This includes many of the habitat-forming corals that create vulnerable marine ecosystems, including scleractinian stony corals, antipatharian black corals, octocorals and stylasterid lace corals, and habitat-forming sponges. The baseline state of CWCS has declined since the second World Ocean Assessment. Fishing in deep waters continues around the world, and oil and gas development is entering new areas and moving deeper in the ocean. Deep-sea mining and marine CO2 removal have both entered the trial phase and have the potential to be stressors in the near future Ref 38. In addition, climate change is a pervasive threat and the deep ocean absorbs a considerable amount of heat and CO2.
Among the most significant recent academic advances is the publication of an edited volume in the Coral Reefs of the World series entitled Cold-Water Coral Reefs of the World Ref 18. This includes descriptive summaries of CWCS ecosystems in various regions of the world, along with a summary chapter establishing a new global model of scleractinian coral habitat suitability, including the suitability of habitats for coral reef formation (see figure I). According to that analysis, cold-water coral reefs form in areas with especially high current speeds and dissolved oxygen availability, as compared with areas where corals are present but do not form large reef structures. Similar approaches have also been used to explore future suitable areas for habitat-forming sponges, highlighting how depth, temperature and dissolved oxygen are key drivers Ref 31.
Over the past few years, the Blake Plateau along the Atlantic coast of the United States of America has been completely mapped using multibeam sonar, which revealed over 30,000 km2 of CWCS mounds, including a continuous area covering 26,000 km2 Ref 64. This revealed new areas of cold-water coral reefs beneath the Gulf Stream that appeared to have some of the most variable oceanographic conditions observed, including temperature swings from 4℃ to over 10℃ in 24 hours, and among the highest nitrate concentrations recorded in the Western Atlantic Ref 17. Studies have also highlighted how CWCS mounds grow into the prevalent directions of the current Ref 15, altering hydrodynamics to facilitate mound growth and sedimentation Ref 66. Paleoecological studies of cold-water coral mounds have revealed some of the key drivers of their persistence through time, demonstrating the importance of food supply and dissolved oxygen concentration Ref 57 Ref 53, which are also affected by large- and small-scale hydrodynamics.
On a finer scale, in the present day, the biomass distribution on CWCS reefs has been found to demonstrate metre-scale fauna clustering on the reef structure Ref 20. Sponges within CWCS reefs can act by recycling dissolved organic matter into particulate detritus, which can then be ingested by other CWCS fauna Ref 4. This "sponge loop" contributes to the overall food availability of CWCS habitats and should be considered in future projections. The corals forming CWCS are well adapted to highly variable food delivery Ref 13 and the communities they support have evolved to optimize carbon and energy transfer through the food web Ref 41. Lophelia pertusa, synonymized as Desmophyllum pertusum, was found to be well adapted to fluctuations in food supply through high capture capacities, resource flexibility, investment in tissue reserves and synchronizing activity to seasonal fluctuations in food ability Ref 42. Recently developed methods have been used to determine that the carbon turnover of cold-water coral reefs may be approximately three times higher than that of soft-sediment areas at similar depths Ref 21. Biogeochemical metabolic flux models of how deep-sea sponges and their microbial communities transform different types of carbon and nitrogen shed light on how dissolved organic matter acts as a primary energy source for sponges Ref 22.
The threat of climate change to CWCS will vary by region and depth and will include changes in temperature, deoxygenation, ocean acidification and reduced food supply. Corals or sponges with narrower environmental ranges will be more at risk Ref 24, and the slow growth rate of these deep-sea organisms, coupled with limited dispersal capabilities in some cases, places them at particular risk Ref 18 Ref 31. The onset of ocean acidification is of particular concern to the dead exposed coral skeleton, leading to the dissolution of the aragonitic skeleton and "coralporosis" Ref 34. This can lead to a loss of structural integrity Ref 71 and the collapse of the coral framework (see figure II). The loss of the dead framework will act directly on the types of biodiversity within CWCS ecosystems as reef sessile suspension feeders cannot attach to strongly degraded reef framework and coral rubble Ref 41. This will, in turn, affect nutrient recycling within the habitat and also change local hydrodynamics, which may affect food capture and the growth of remaining live coral Ref 15.
Restoration activities are increasing, leading to greater understanding of the associated techniques and challenges for both framework- and non-framework-forming coral species. This includes guides on techniques Ref 7, reviews of the challenges to overcome with recommendations for future work Ref 39 Ref 46 Ref 10 and advice on how local communities and stakeholders can be integrated into restoration efforts Ref 47. There is at present no overarching global scheme for the conservation and restoration of CWCS ecosystems, although they are generally considered vulnerable marine ecosystems in the European parlance and managed as essential fish habitat in the United States.
2. Environmental changes since the second World Ocean Assessment
While advances have been made in understanding of environmental changes since the second World Ocean Assessment, a lack of long-term baseline data at CWCS sites still limits understanding of long-term change. It is clear that the deep sea is warming (well established), but it is difficult to say how quickly (inconclusive). There is evidence of more frequent "marine heatwaves" (well established) but less information on how deep they penetrate and how frequently they reach the deep seafloor (unresolved). The risks and potential impacts of typical offshore industrial activities Ref 16 and catastrophic incidents, such as the Deepwater Horizon disaster, are well established Ref 62. This has not curtailed offshore drilling, however. Similarly, for fisheries, the impacts of bottom trawling on benthic communities are understood (well established), including as regards CWCS Ref 25 Ref 70.
To understand the threat of climate change to CWCS ecosystems, there is a need to combine both experimental and in situ studies and observations. This combination has been used in environmental niche modelling to predict changes in habitat suitability for key cold-water coral species, with declines of between 28% and 100% predicted by the end of the century in high-emission scenarios (established but incomplete). The largest reductions were for the framework-forming Desmophylum pertusum and the octocoral Paragorgia arborea Ref 48. According to predicted distribution models, the trend for sponges will be similar to that of cold-water corals: habitats may shift (and expand, in some cases, with the area suitable for the glass sponge Vazella pourtalesii on the Scotian Shelf increasing by up to four times its current size) but there will be an overall trend of reduced suitability in areas where these habitats are currently found Ref 6. An added complication is the fact that the impacts of climate change will take effect at the same time as those of established human stressors (such as fishing) and emerging human stressors (such as mining and marine CO2 removal). For example, bottom trawling at depths in excess of 800 m was banned in Europe in 2016, but these activities continue in many other areas of the world.
Figure I Map of ensemble weighted mean habitat suitability index scores for CWCS reefs and reef-forming taxa known to occur within the regions of the world

There is a lack of data on how long it may take for cold-water coral reefs to be affected by climate change. Paleo-records of key framework-forming species are of great benefit in this regard (Portilho- Ramos, 2022), as observation of impacts in situ is limited by regional capacity for monitoring and repeated observation over potentially decadal timescales. Paleo-records can also be used to identify key drivers of past impacts (e.g. food supply through local hydrodynamics) and provide insights into how present-day reefs can survive in sub-optimal conditions (e.g. low oxygen), such as CWCS off the Mauritanian Margin Ref 53. There have been very few studies of the effects of climate change on coral gardens (consisting of octocoral sea fans, antipatharian black corals, stylasterid hydrocorals and others) and sponge beds (unresolved).
Figure II Cold-water coral habitat loss in a future ocean
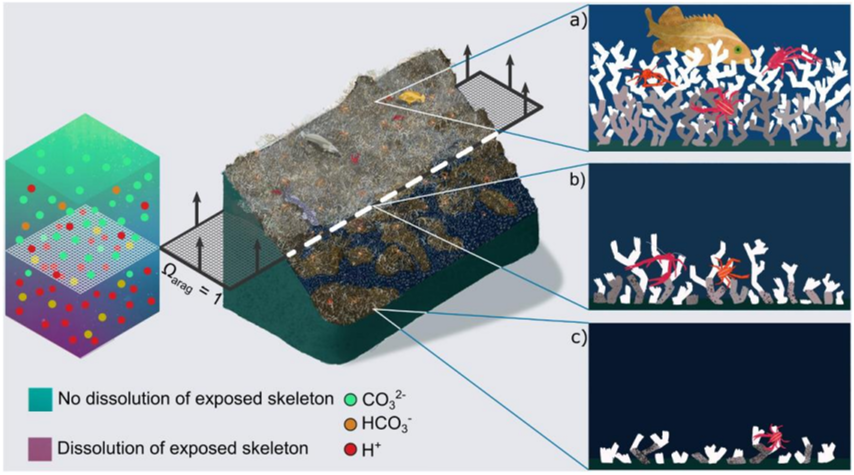
Note: The central image shows Desmophyllum pertusum reefs under present-day conditions (top) and their projected habitat loss in a future ocean once the aragonite saturation horizon (dashed layer at Q2Arag = 1) rises above them. A qualitative chemistry scale bar indicates dissolution of exposed aragonite beneath the horizon driven by increased [H+] and decreased [CO32-] ions. Arrows on the dashed layer indicate the projected direction of movement of the aragonite saturation horizon driven by ocean acidification. Panels on the right zoom into areas of present-day and future reefs related to the position of the aragonite saturation horizon. In (a), a healthy reef is shown with a substantial live (white) and dead (grey) coral framework. In (b), porosity has started to appear in the dead, exposed skeletal framework, leading to crumbling. In (c), the exposed skeletal framework exhibits large porosity and is unable to support a complex three-dimensional habitat, although high abundances of live coral can still exist.
3. Region-specific changes
Arctic Ocean
The number of studies of CWCS, which are pan-Arctic ecosystems, has increased since 2018 with the aim of identifying key biodiversity hotspots of CWCS across several areas, including the continental shelves of Greenland and Norway, as well as fjords around Svalbard Ref 55. The Davis Strait hosts important sponge grounds and soft coral fields Ref 40, but the inaccessibility of some areas (due to such factors as sea ice) means that these studies are often based on singular or episodic observations. The Karasik seamount expedition revealed that some deep-sea sponges, such as Geodia parva and Stelletta rhaphidiophora, can move across the seafloor Ref 50.
North Atlantic Ocean, Baltic Sea, Black Sea, Mediterranean and North Sea
Understanding of how overturning circulations provide surface food supply to reefs around the North and Eastern Atlantic has improved Ref 35 Ref 23, including understanding of historical patterns in the Baltic Sea Ref 57. In particular, it is clear that current CWCS reefs may be at risk due to water column stratification associated with climate change. Quantification of water mass dynamics also supports better understanding of larval flow between CWCS areas Ref 61 and the identification of unique sponge fauna in the Mediterranean Ref 59. Since 2019, there has been some progress in the protection of cold-water coral habitat in such areas as Eastern Canyons Marine Refuge in Canada, the West of Scotland marine protected area (MPA) and the Tristan da Cunha MPA in the Southern Atlantic.
South Atlantic Ocean and wider Caribbean
Despite the dire need to curb fossil fuel use, offshore drilling continues to expand around the world, moving into frontier areas of the deep ocean in the southern Caribbean, West Africa and South America. Detailed mapping of the mounds off Brazil Ref 58 and Mauritania has revealed a wider distribution and greater abundance of CWCS than previously thought. The systems off West Africa seem to be surviving, and in some cases thriving in waters with extremely low oxygen, in contrast to findings elsewhere Ref 33. New taxonomic and coral distribution work in South Africa Ref 27 is strengthening the knowledge base with the discovery of many coral gardens, and new offshore MPAs have improved CWCS habitat protection, with eco-certification initiatives for fisheries supporting further improvements in the management of CWCS and vulnerable marine ecosystems Ref 63.
Indian Ocean, Arabian Sea, Bay of Bengal, Red Sea, Gulf of Aden and Persian Gulf
Taxonomic knowledge of CWCS is advancing in Africa Ref 27 Ref 32 Ref 60 and museum records are being used to examine diversity patterns in the Indian and Atlantic Oceans Ref 28. There has been little progress in mapping cold-water coral habitats along the African continental margin, but new expeditions in the Red Sea have increased cold-water coral species and reef discovery and baseline data in that region Ref 56 Ref 14 Ref 43. Cold-water coral protection is lacking along most of the African margin for both the Indian Ocean and the Atlantic Ocean, and there has been limited or no progress in protecting cold-water coral habitats in the high seas of that region. Offshore drilling continues to expand in the Bay of Bengal, Indonesia and elsewhere and continues to threaten CWCS ecosystems before they are even discovered.
North Pacific Ocean
In the Northwestern Hawaiian Islands, gradients in fishery pressure were related to the size and density of corals on seamounts Ref 2. There had been some recovery of corals on seamounts where fishing had occurred historically but not since the 1970s, and which were inside MPAs. The CWCS ecosystems of the Eastern Tropical Pacific have been investigated primarily in Costa Rica and Ecuador Ref 19. Near the Galapagos Islands, large deep-sea coral reefs (at depths of between 400 m and 600 m) were discovered recently. All studies have been single observations, which makes evaluations of change almost impossible. In addition, CWCS have recently been surveyed in the South China Sea, thereby filling a data gap that previously existed in this region Ref 65.
South Pacific Ocean
In addition to long-standing industrial activity, the commercial exploitation of deep-sea mineral resources is an additional threat to CWCS habitats and coral gardens in particular. This is of particular concern in the Central Pacific where most interest has been focused. The potential impacts of mining involve physical destruction of the seabed and damage to the habitat and fauna by mining equipment, as well as changes in seafloor topography and geochemical characteristics in the direct vicinity of the mining site Ref 12. The sediment plumes that are generated can affect the entire water column, disperse for hundreds of kilometres and last several years Ref 51 Ref 49. The extent and nature of ecosystem impacts resulting from these plumes are detrimental, with mortality and tissue loss observed in the octocoral Dentomuricea aff. meteor (Carreiro- Silva and others, 2022). At present, however, combined effects of these impacts and the impacts of environmental change are poorly understood Ref 67.
Southern Ocean
There has been little exploration of CWCS systems in the Southern Ocean in recent years. It should be noted, however, that the presence of the Schmidt Ocean Institute RV Falkor (too) will add significant amounts of data from Antarctica, the South Sandwich Islands and other parts of the area of application of the Convention for the Conservation of Antarctic Marine Living Resources (CCAMLR) in 2024 and 2025. Proposals for additional MPAs in East Antarctica, the Weddell Sea and the Antarctic Peninsula have been under negotiation within the Commission for the Conservation of Antarctic Marine Living Resources but consensus on implementation has not yet been reached Ref 9. The Commission developed binding measures for the identification, designation, monitoring and management of fragile benthic ecosystems, including CWCS, in 2007. These ecosystems have been included in the subsequent design of MPAs throughout the area of application of CCAMLR. In addition, the prohibition of high seas bottom trawling and the targeting of toothfish at depths shallower than 550 m, which entered into force in 2009, closed much of the continental shelf to impacts from bottom contact from fisheries in an effort to avoid significant adverse impacts on CWCS.
4. Key remaining knowledge and capacity gaps and new gaps
Global awareness of the importance of CWCS continues to grow with discoveries of new ecosystems, the services and biodiversity they support and the threats they are under. Research on ecosystem services, climate risks and foundational biodiversity science is being used increasingly in communication efforts. Programmes such as the Deep-Ocean Stewardship Initiative, REV Ocean, OceanX and Ocean Census, as well as the work of the Ocean Exploration Trust with its vessel EV Nautilus and the Schmidt Ocean Institute with its vessel RV Falkor (too), have led to increased international expeditions and philanthropic investment, offshore live streaming and increased focus, training and multimedia science communication efforts (art, animation, documentaries, podcasts, etc.), which are helping to increase access and global awareness.
While awareness of CWCS is growing, the deep sea is coming to be seen as a resource for marine CO2 removal. A number of proposed marine CO2 removal projects entail storing carbon in the deep sea, but there is a danger in overlooking the ecological complexity of the deep sea or failing to account for the habitats and ecosystem processes that may be affected, including CWCS Ref 38. For example, alkalinity enhancement, direct injection of CO2 or organic carbon deposition could directly or indirectly alter local pH, oxygen levels and sedimentation dynamics, and CWCS, including their larval stages, are potentially sensitive to smothering from an excess of organic matter or mineral deposition (see Levin and others (2023) for a review of these issues).
Looking forward, the conservation and sustainable management of CWCS aligns with the Sustainable Development Goals, particularly Goal 14: Conserve and sustainably use the oceans, seas and marine resources for sustainable development. Ecosystem services from CWCS habitats are at risk, including climate regulation, food production and the provision of livelihoods Ref 48 Ref 17. CWCS also play an indirect and important role in carbon sequestration over the intermediate-term (hundreds or thousands of years) by provisioning their complex skeletons and building the habitats that support a diverse range of marine life, which all contribute to the deep-sea carbon cycle Ref 5.
Integrating CWCS into climate adaptation and mitigation policies, through nationally determined contributions, for instance, would aid their role in supporting biodiversity, maintaining ecosystem services and contributing to global decarbonization goals. For example, aligning the Antarctic Treaty System with the Paris Agreement could facilitate the inclusion of Antarctic CWCS conservation targets in nationally determined contributions Ref 5. Morato and others (2020) emphasize the need for better integration of climate refugia into spatial management efforts. There have also been improvements in terms of accounting for the ecosystem services provided by cold-water coral habitat Ref 17 and new work to support consideration of these services in climate-smart marine spatial planning Ref 10 Ref 37. This supports ecosystem service-based management.
There is also increasing recognition of the importance of Indigenous knowledge and appreciation for cultural values in understanding and managing deep-sea ecosystems Ref 26. Cultural connections to the deep sea include spiritual connections, and Indigenous coastal communities are expressing increased concern for deep-sea habitats, including CWCS. Policy briefs and arts-based approaches have offered new avenues for understanding, documenting and sharing traditional and Indigenous knowledge Ref 26.
Capacity for monitoring and research is overwhelmingly concentrated in the global North. Most countries lack deep-water monitoring and many lack research vessels, resulting in a dependence on outside technologies and data-sharing. The FAIR data principles (which state that data should be Findable, Accessible, Interoperable and Reusable) are essential for promoting open and equitable data-sharing, and they are complemented by the Collective Benefit, Authority to Control, Responsibility, Ethics framework, under which data collection and sharing should be conducted ethically, inclusively and with respect for the rights and knowledge of local communities. The Nagoya Protocol on Access to Genetic Resources and the Fair and Equitable Sharing of Benefits Arising from Their Utilization to the Convention on Biological Diversity provides an important framework to ensure that the benefits arising from samples and data are shared with local communities on a fair and equitable basis.
To support growing observation efforts, technologies that enhance monitoring and protection for management increasingly include autonomous systems (Zelada-Leon and others, 2020). These advances support the development of indicators for assessing key deep-sea habitats in national waters and areas beyond national jurisdiction, and effectively reduce habitat disturbance by improving marine spatial planning Ref 36 Ref 52. Environmental DNA technologies are rapidly advancing and could enable low-cost, non-invasive, efficient monitoring of biodiversity and ecosystem health Ref 44, especially when combined with curated sequence libraries and taxonomic reference collections. In addition to environmental DNA, recent advancements in technology, including crewed submersibles and artificial intelligence, have also significantly enhanced understanding of CWCS. While these tools still face certain limitations, these technologies enable deeper and more detailed exploration, allowing for in situ observations of growth, behaviour and species interactions Ref 45 Ref 54 Ref 1 Ref 29 Ref 69. These advances in technology and data acquisition, in combination with early detection of stressors (such as pollution, temperature changes and invasive species), enables faster management responses, reducing damage to CWCS and potentially preventing habitat degradation.
References
- Aguzzi, J., Thomsen, L., Flögel, S., Robinson, N.J., Picardi, G., Chatzievangelou, D., and others (2024). New technologies for monitoring and upscaling marine ecosystem restoration in deep-sea environments. Engineering 34, 195-211.
- Baco, A.R., Morgan, N.B., Roark, E.B., Biede, V. (2023). Bottom-contact fisheries disturbance and signs of recovery of precious corals in the Northwestern Hawaiian Islands and Emperor Seamount Chain. Ecological Indicators, 148:110010.
- Barnhill, K.A., Roberts, J.M., Myers-Smith, I., and others (2023). Incorporating dead material in ecosystem assessments and projections. Nat. Clim. Chang. 13, 113-115. https://doi.org/10.1038/s41558- 022-01565-5.
- Bart, M.C., Hudspith, M., Rapp, H.T., Verdonschot, P.F.M., and de Goeij, J.M. (2021). A Deep-Sea Sponge Loop? Sponges Transfer Dissolved and Particulate Organic Carbon and Nitrogen to Associated Fauna. Front. Mar. Sci., 8:604879. doi: 10.3389/fmars.2021.604879.
- Bax, N., Sands, C.J., Gogarty, B., Downey, R.V., Moreau, C.V., Moreno, B., Held, C., Paulsen, M.L., McGee, J., Haward, M. and Barnes, D.K., (2021). Perspective: increasing blue carbon around Antarctica is an ecosystem service of considerable societal and economic value worth protecting. Global Change Biology, 27(1), pp.5-12.
- Beazley, L., Kenchington, E., Murillo, F.J., Brickman, D., Wang, Z., Davies, A.J., Roberts, E.M., Rapp, H.T. (2021). Climate change winner in the deep sea? Predicting the impacts of climate change on the distribution of the glass sponge Vazella pourtalesii. Marine Ecology Progress Series, 657:1-23. https://doi.org/10.3354/meps13566.
- Boch, C.A., A. De Vogelaere, E.J. Burton, C. King, C. Lovera, K. Buck, J. Lord, L. Kuhnz, M. Kaiser, C. Reid-Rose, and J.P. Barry. (2020). Guide to translocating coral fragments for deep-sea restoration. National Marine Sanctuaries Conservation Series ONMS-20-10. U.S. Department of Commerce, National
- Oceanic and Atmospheric Administration, Office of National Marine Sanctuaries, Silver Spring, MD. 25 pp.
- Boothroyd, A., Adams, V., Alexander, K., and Hill, N. (2024). Stakeholder perceptions of the Commission for the Conservation of Antarctic marine living resources marine protected area planning process. Marine Policy, 170, 106381.
- Bravo, M.E., Brandt, M.I., Van Der Grient, J.M.A., Dahlgren, T.G., Garrote, P.E., Gollner, S., Jones, D.O.B., Levin, L.A., McClain, C.R., Narayanaswamy, B.E., Sutton, T.T., Vicorero, L., Cordes, E.E. (2023). Insights from the management of offshore energy resources: Towards an ecosystem-services based management approach for deep-ocean industries. Front Mar Sci., 9: 994632. doi: 10.3389/ fmars.2022.994632.
- Büscher, J.V., Juva, K., Flögel, S., Wisshak, M., Rüggeberg, A., Riebesell, U., Form, A.U. (2024). Water mass characteristics and hydrodynamics at an inshore versus an offshore mid-Norwegian cold-water coral reef habitat. Front. Mar. Sci., 11:1363542. https://doi.org/10.3389/fmars.2024.1363542.
- Carreiro-Silva, M., Martins, I., Riou, V., Raimundo, J., Caetano, M., Bettencourt, R., and others (2022). Mechanical and toxicological effects of deep-sea mining sediment plumes on a habitat-forming cold- water octocoral. Frontiers in Marine Science, 9:915650. https://doi.org/10.3389/fmars.2022.915650.
- Carrick, J.V., Mienis, F., Rossby, T., Cordes, E.E., Demopoulos, A.M. and Davies, A.J. (2024). Gulf Stream intrusion and deep current upwelling drive dynamic patterns of temperature and food supply within cold-water coral reefs. Limnology and Oceanography, 69: 2193-2210. doi: 10.1002/lno.12659.
- Chimienti, G., Terraneo, T.I., Vicario, S., Marchese, F., Purkis, S.J., Eweida, A.A., Rodrigue, M., and Benzoni, F. (2022). A new species of Bathypathes (Cnidaria, Anthozoa, Antipatharia, Schizopathidae) from the Red Sea and its phylogenetic position. ZooKeys, 1116, 1-22. https://doi.org/10.3897/zookeys.1116.79846.
- Corbera, G., Lo Iacono, C., Simarro, G., and others (2022). Local-scale feedbacks influencing cold-water coral growth and subsequent reef formation. Sci Rep, 12, 20389. https://doi.org/10.1038/s41598-022- 24711-7.
- Cordes, E.E., Jones, D.O.B., Schlacher, T.A., Amon, D.J., Bernardino, A.F., Brooke, S., Carney, R., DeLeo, D.M., Dunlop, K.M., Escobar, E., Gates, A.R., Génio, L., Gobin, J., Henry, L-A., Herrera, S., Hoyt, S., Joye, S., Kark, S., Mestre, N.C., Metaxas, A., Pfeifer, S., Sink, K., Sweetman, A.K., Witte, U. (2016). Environmental impacts of the deep-water oil and gas industry: a review to guide management strategies. Front Environ Sci, 4: 58. doi: 10.3389/fenvs.2016.00058.
- Cordes, E.E., Demopoulos, A.W.J., Davies, A.J., and others (2023). Expanding our view of the cold-water coral niche and accounting of the ecosystem services of the reef habitat. Sci Rep, 13, 19482. https://doi.org/10.1038/s41598-023-45559-5.
- Cordes, E.E., Mienis (2024). The Cold-Water Coral Reefs of the World. Volume 19: Coral Reefs of the World series. Springer International Publishing. doi: 10.1007/978-3-031-40897-7.
- Cortés, J. (2019). Chapter 26: Isla del Coco, Costa Rica, Eastern Tropical Pacific, pp. 465-474. In Y. Loya, K.A. Puglise, T.C.L. Bridge, eds. Mesophotic Coral Ecosystems. Coral Reefs of the World 12. Springer Nature, Switzerland. https://doi.org/10.1007/978-3-319-92735-0_26.
- De Clippele, L.H., Rovelli, L., Ramiro-Sánchez, B., Kazanidis, G., Vad, J., Turner, S., Glud, R.N., and Roberts, J.M. (2021a). Mapping cold-water coral biomass: an approach to derive ecosystem functions. Coral Reefs 40, 215-231.
- De Clippele, L.H., van der Kaaden, A .- S., Maier, S.R., de Froe, E., and Roberts, J.M. (2021b). Biomass mapping for an improved understanding of the contribution of cold-water coral carbonate mounds to C and N cycling. Frontiers in Marine Science, 8, 1608.
- De Kluijver ,A., Bart M.C., van Oevelen, D., de Goeij, J.M., Leys, S.P., Maier, S.R., Maldonado, M., Soetaert, K., Verbiest, S., and Middelburg, J.J. (2021). An integrative model of carbon and nitrogen metabolism in a common deep-sea sponge (Geodia barretti). Front. Mar. Sci., 7, 596251. https://doi.org/10.3389/fmars.2020.596251.
- De Froe, E., Maier, S.R., Horn, H.G., Wolff, G.A., Blackbird, S., Mohn, C., Schultz, M., van der Kaaden, A .- S., Cheng, C.H., Wubben, E., van Haastregt, B., Moller, E.F., Lavaleye, M., Soetaert, K., Reichart, G .- J., and others (2022). Hydrography and food distribution during a tidal cycle above a cold-water coral mound. Deep-Sea Research I 189, 103854. https://doi.org/10.1016/j.dsr.2022.103854.
- Dijkstra, J.A., Mello, K., Sowers, D., Malik, M., Watling, L., and Mayer, L.A. (2021). Fine-scale mapping of deep-sea habitat-forming species densities reveals taxonomic specific environmental drivers. Global Ecology and Biogeography, 30(6), 1286-1298.
- Du Preez, C., Swan, K.D., Curtis, J.M. (2020). Cold-water corals and other vulnerable biological structures on a North Pacific seamount after half a century of fishing. Frontiers in Marine Science, 7:17.
- Escobar, E., B. Guilloux, K. Howell, D. Jones, C. Koyavakauta, L. Levin, C.Y. Mulalap, P. Singh, K. Sink, V. Tilot, P. Turner, A. Vencatesan, K. Willaert (2021). The necessity of traditional knowledge for management of deep-seabed mining. Deep-Ocean Stewardship Initiative.
- Filander, Z.N., Kitahara, M.V., Cairns, S.D., Sink, K.J., and Lombard, A.T. (2021). Azooxanthellate Scleractinia (Cnidaria, Anthozoa) from South Africa. ZooKeys, 1066, 1-198. https://doi.org/10.3897/zookeys.1066.69697.
- Filander, Z.N., Sink, K.J., Kitahara, M.V., Cairns, S.D., Lombard, A.T. (2024). Diversity patterns of the South African azooxanthellate scleractinians (Cnidaria: Anthozoa), with considerations of environmental correlates. PLoS ONE, 19(8): e0296188. https://doi.org/10.1371/journal.pone.0296188.
- Gallego, R., Arias, M.B., Corral-Lou, A., Díez-Vives, C., Neave, E.F., Wang, C., and Riesgo, A. (2024). North Atlantic deep-sea benthic biodiversity unveiled through sponge natural sampler DNA. Communications Biology, 7(1), 1015.
- Gómez, C.E., Gori, A., Weinnig, A.M., Hallaj, A., Chung, H.J. and Cordes, E.E. (2022) Natural variability in seawater temperature compromises the metabolic performance of a reef-forming cold-water coral with implications for vulnerability to ongoing global change. Coral Reefs. 41: 1225-1237. doi: 10.1007/s00338-022-02267-2.
- Gregório, I., Xavier, J.R., and Davies, A.J. (2024). Present and future distribution of the deep-sea habitat- forming sponge-Pheronema carpenteri () in a changing ocean. Deep Sea Research Part I: Oceanographic Research Papers, 213, 104390.
- Hanafi-Portier, M., Samadi, S., Corbari, L., Boulard, M., Miramontes, E., Penven, P., Leroy, B., Napoléon, T., Jorry, S.J. and Olu, K. (2024). Multiscale spatial patterns and environmental drivers of seamount and island slope megafaunal assemblages along the Mozambique channel. Deep Sea Research Part I: Oceanographic Research Papers, 203, 104198. https://doi.org/10.1016/j.dsr.2023.104198
- Hebbeln, D., Portilho-Ramos, R.D., Wienberg, C., Titschack, J. (2019). The fate of cold-water corals in a changing world: a geological perspective. Frontiers in Marine Science, 6:119.
- Hennige, S.J., Wolfram, U., Wickes, L., Murray, F., Roberts, J.M., Kamenos, N.A., Schofield, S., Groetsch, A., Spiesz, E.M., Aubin-Tam, M.E., Etnoyer, P.J. (2020). Crumbling reefs and cold-water coral habitat loss in a future ocean: evidence of "Coralporosis" as an indicator of habitat integrity. Frontiers in Marine Science, 7, 668. https://doi.org/10.3389/fmars.2020.00668.
- Juva, K., Flögel, S., Karstensen, J., Linke, P., and Dullo, W-C. (2020). Tidal dynamics control on cold- water coral growth: A high-resolution multivariable study on Eastern Atlantic cold-water coral sites. Front. Mar. Sci., 7:132. https://doi.org/10.3389/fmars.2020.00132.
- Kazanidis, G., Orejas, C., Borja, A., Kenchington, E., Henry, L-A., Callery, O., Carreiro-Silva, M., Egilsdottir, H., Giacomello, E., Grehan, A., Menot, L., Morato, T., Ragnarsson, S.A., Rueda, J.L., Stirling, D., Stratmann, T., van Oevelen, D., Palialexis, A., Johnson, D., Roberts, J.M. (2020). Assessing the environmental status of selected North Atlantic deep-sea ecosystems. Ecological Indicators, 119: 106624.
- La Bianca, G., Rees, S., Attrill, M.J., Lombard, A.T., McQuaid, K.A., Niner, H.J., van Rein, H., Sink, K.J., Howell, K.L. (2023). A standardised ecosystem services framework for the deep sea. Frontiers in Marine Science, 10: 1176230.
- Levin, L., Alfaro-Lucas, J., Colaço, A., Cordes, E.E., Craik, N., Danovaro, R., Hoving, H-J., Ingels, J., Mestre, N., Seabrook, S., Thurber, A., Vivian, C., Yasuhara, M. (2023). Deep-sea impacts of climate interventions. Science, 379: 978-981. doi: 10.1126/science.ade7521.
- Liu, and others (2024). CORDAP RandD roadmap for conservation and restoration of cold-water corals. CORDAP. www.cordap.org.
- Maier, S.R., Arboe, N.H., Christiansen, H., Krawczyk, D.W., Meire, L., Mortensen, J., Planken, K., Schulz, K., van der Kaaden, A.S., Vonnahme, T.R., Zwerschke, N. (2024). Arctic benthos in the Anthropocene: Distribution and drivers of epifauna in West Greenland. Science of the Total Environment, 951:175001.
- Maier, F.M., E. de Froe, K. Soetaert, M. Lavaleye, G. Duineveld, O. Beauchard, A .- S. van der Kaaden, B.P. Koch, D. van Oevelen (2021). Reef communities associated with 'dead' cold-water coral framework drive resource retention and recycling in the deep sea. Deep Sea Research Part I: Oceanographic Research Papers, 75, 103574. https://doi.org/10.1016/j.dsr.2021.103574.
- Maier, S.R., Brooke, S., De Clippele, L.H., de Froe, E., van der Kaaden, A .- S., Kutti, T., Mienis, F., van Oevelen, D. (2023). On the paradox of thriving cold-water coral reefs in the food-limited deep sea. Biol Rev, 98: 1768-1795. https://doi.org/10.1111/brv.12976.
- Maggioni, D., Terraneo, T.I., Chimienti, G., Marchese, F., Pica, D., Cairns, S.D., Eweida, A.A., Rodrigue, M., Purkis, S.J., and Benzoni, F. (2022). The first deep-sea stylasterid (Hydrozoa, Stylasteridae) of the Red Sea. Diversity, 14 (4), 241. https://doi.org/10.3390/d14040241.
- McCartin, L.J., Voshen, S.A., Ambrose, S.W., Layden, M., McFadden, C.S., Cordes, E.E., McDermott, J.M., Herrera, S. (2022). Temperature controls eDNA persistence across physiochemical conditions in seawater. ES&T, 56: 8629-8639. doi: 10.1021/acs.est.2c01672.
- Mogstad, A.A., Løvås, H.S., Sture, Ø., Johnsen, G., and Ludvigsen, M. (2022). Remote sensing of the Tautra Ridge: an overview of the World's shallowest cold-water Coral Reefs. Frontiers in Marine Science, 9, 848888.
- Montseny, M., Linares, C., Carreiro-Silva, M., Henry, L .- A., Billett, D., Cordes, E.E., Smith, C.J., Papadopoulou, N., Bilan, M., Girard, F., Burdett, H.L., Larsson, A., Strömberg, S., Viladrich, N., Barry, J.P., Baena, P., Godinho, A., Grinyó, J., Santín, A., and Gori, A. (2021a). Active Ecological Restoration of Cold-Water Corals: Techniques, Challenges, Costs and Future Directions. Frontiers in Marine Science, 8, Article 621151. https://doi.org/10.3389/fmars.2021.621151.
- Montseny, M., Linares, C., Viladrich, N., Biel, M., Gracias, N., Baena, P., Quintanilla, E., Ambroso, S., Grinyó, J., Santín, A. and Salazar, J., (2021b). Involving fishers in scaling up the restoration of cold-water coral gardens on the Mediterranean continental shelf. Biological conservation, 262, p.109301.
- Morato ,T., J.M. González-Irusta, C. Dominguez-Carrió, C.L. Wei, A. Davies, A.K. Sweetman, and others (2020). Climate-induced changes in the suitable habitat of cold-water corals and commercially important deep-sea fishes in the North Atlantic. Glob Chang Biol., 26(4), 2181-2202. https://doi.org/10.1111/gcb.14996.
- Morato, T., Juliano, M., Pham, C.K., Carreiro-Silva, M., Martins, I., and Colaço A. (2022). Modelling the dispersion of Seafloor Massive Sulphide mining plumes in the Mid Atlantic Ridge around the Azores. Frontiers in Marine Science, 9. https://doi.org/10.3389/fmars.2022.910940.
- Morganti, T.M., Slaby, B.M., de Kluijver, A., Busch, K., Hentschel, U., Middelburg, J.J., Grotheer, H., Mollenhauer, G., Dannheim, J., Rapp, H.T., Purser, A. (2022). Giant sponge grounds of Central Arctic seamounts are associated with extinct seep life. Nature communications, 13:638.
- Muñoz-Royo, C., Peacock, T., Alford, M.H., Smith, J.A., Le Boyer, A., Kulkarni, C.S., and others (2021). Extent of impact of deep-sea nodule mining midwater plumes is influenced by sediment loading, turbulence and thresholds. Communications Earth & Environment, 2, 148.
- Orejas, C., Kenchington, E., Rice, J., Kazanidis, G., Palialexis, A., Johnson, D., Gianni, M., Danovaro, R., Roberts, J.M. (2020). Towards a common approach to the assessment of the environmental status of deep- sea ecosystems in areas beyond national jurisdiction. Marine Policy, 121, 104182. https://doi.org/10.1016/j.marpol.2020.104182.
- Portilho-Ramos, R.D., Titschack, J., Wienberg, C., Siccha Rojas, M.G., Yokoyama, Y., and Hebbeln, D. (2022). Major environmental drivers determining life and death of cold-water corals through time. PLoS BIOLOGY, 20(5), e3001628. https://doi.org/10.1371/journal.pbio.3001628.
- Prado, E., Abad-Uribarren, A., Ramo, R., Sierra, S., González-Pola, C., Cristobo, J., and Sánchez, F. (2023). Describing polyps behavior of a deep-sea gorgonian, Placogorgia sp., using a deep-learning approach. Remote Sensing, 15(11), 2777.
- Puerta, P., Johnson, C., Carreiro-Silva, M., Henry, L.A., Kenchington, E., Morato, T., Kazanidis, G., Rueda, J.L., Urra, J., Ross, S., Wei, C.L. (2020). Influence of water masses on the biodiversity and biogeography of deep-sea benthic ecosystems in the North Atlantic. Frontiers in Marine Science, 7:239.
- Qurban, M.A., Krishnakumar, P.K., Joydas, T.V., and others (2020). Discovery of deep-water coral frameworks in the northern Red Sea waters of Saudi Arabia. Sci Rep, 10, 15356. https://doi.org/10.1038/s41598-020-72344-5.
- Raddatz, J., Liebetrau, V., Rüggeberg, A., and others (2020a). Living on the edge: Environmental variability of a shallow late Holocene cold-water coral mound. Coral Reefs, 41, 1255-1271. https://doi.org/10.1007/s00338-022-02249-4.
- Raddatz, J., Titschack, J., Frank, N., Freiwald A, Conforti A, Osborne A, Skornitzke S, Stiller W, Rüggeberg A, Voigt S, Albuquerque AL. (2020b). Solenosmilia variabilis-bearing cold-water coral mounds off Brazil. Coral Reefs 39, 69-83.
- Santín, A., Grinyó, J., Uriz, M.J., Lo Iacono, C., Gili, J.M., and Puig, P. (2021). Mediterranean Coral Provinces as a Sponge Diversity Reservoir: Is There a Mediterranean Cold-Water Coral Sponge Fauna? Front. Mar. Sci., 8, 662899. https://doi.org/10.3389/fmars.2021.662899.
- Samaai, T., Kelly, M., Ngwakum, B., Payne, R., Teske, P.R., Janson, L., Kerwath, S., Parker, D., Gibbons, M.J. (2020). New Latrunculiidae (Demospongiae, Poecilosclerida) from the Agulhas ecoregion of temperate southern Africa. Zootaxa, 4896(3). https://doi.org/10.11646/zootaxa.4896.3.4.
- Schulz, K., Soetaert, K., Mohn, C., Korte, L., Mienis, F., Duineveld, G., van Oevelen, D. (2020). Linking large-scale circulation patterns to the distribution of cold water corals along the eastern Rockall Bank (northeast Atlantic). Journal of Marine Systems, 212, 103456.
- Schwing, P., Montagna, P.A., Joye, S.B., Paris, C.B., Cordes, E.E., McClain, C.R., Kilborn, J.P., Murawski, S.A. (2020). Deep benthic ecosystem impacts of the Deepwater Horizon oil spill: Assembling the record of species and community change. Front Mar Sci., 7:560012. doi: 10.3389/ fmars.2020.560012.
- Sink, K.J., Lombard, A.T., Attwood, C.G., Livingstone, T.C., Grantham, H., Holness, S.D. (2023). Integrated systematic planning and adaptive stakeholder process support a 10-fold increase in South Africa's Marine Protected Area estate. Conservation Letters, 16:e12954.
- Sowers, D.C., Mayer, L.A., Masetti, G., Cordes, E.E., Gasbarro, R., Lobecker, E., Cantwell, K., Candio, S., Hoy, S., Malik, M., White, M., Dornback, M. (2024) Mapping and Geomorphic Characterization of the Vast Cold-Water Coral Mounds of the Blake Plateau. Geomatics 4: 17-47. doi: 10.3390/ geomatics4010002.
- Ta, K., Song, X., Wei, Z., Du, M., Xu, H., Chen, S., and Peng, X. (2024). Oasis of the deep: Cold-water corals of the South China Sea. Marine Environmental Research, 195, 106354.
- Van der Kaaden, A.S., Van Oevelen, D., Rietkerk, M., Soetaert, K., Van de Koppel, J. (2020). Spatial self- organization as a new perspective on cold-water coral mound development. Frontiers in Marine Science, 7,631.
- Van Dover, C.L., Ardron, J.A., Escobar, E., Gianni, M., Gjerde, K.M., Jaeckel, A., and others (2017). Biodiversity loss from deep-sea mining. Nature Geoscience, 10(7), 464-465.
- Van Dover, C.L., Colaço, A., Collins, P.C., Croot, P., Metaxas, A., Murton, B.J., and others (2020). Research is needed to inform environmental management of hydrothermally inactive and extinct polymetallic sulfide (PMS) deposits. Marine Policy, 121, 104183.
- Wang, W., Sun, Y. F., Gao, W., Xu, W., Zhang, Y., and Huang, D. (2024). Quantitative detection algorithm for deep-sea megabenthic organisms based on improved YOLOv5. Frontiers in Marine Science, 11, 1301024.
- Williams, A., Althaus, F., Maguire, K., Green, M., Untiedt, C., Alderslade, P., Clark, M.R., Bax, N., Schlacher, T.A. (2020). The fate of deep-sea coral reefs on seamounts in a fishery-seascape: what are the impacts, what remains, and what is protected ?. Frontiers in Marine Science, September 25, 7:567002.
- Wolfram, U., Peña Fernández, M., McPhee, S., Smith, E., Beck, R.J., Shephard, J.D., Ozel, A., Erskine, C.S., Büscher, J., Titschack, J., Roberts, J.M. (2022). Multiscale mechanical consequences of ocean acidification for cold-water corals. Scientific Reports, 12: 8052.
- Zelada Leon, A., Huvenne, V.A., Benoist, N.M., Ferguson, M., Bett, B.J., Wynn, R.B. (2020). Assessing the repeatability of automated seafloor classification algorithms, with application in marine protected area monitoring. Remote Sensing, 12:1572.