Pelagic domain
Writing team: Marta Coll (coordinating author), Camila Indira Artana, José María Bellido, Peter Croot, Antonio Di Natale, Flavia Lucena Frédou, Susana Garrido, Rafael González-Quirós (lead member), Ryan Henegan, Brian Hunt, Francisco Ramírez, James J. Ruzicka, Lynne J. Shannon, Lloyd Spencer Davis, Akinori Takasuka and Tymon Zielinski (co-lead member).
Key points
The pelagic domain encompasses the entire water column and is the largest oceanic region. Due to its vastness and limited accessibility, it remains less studied and understood than its benthic and intertidal counterparts. The existing knowledge underscores its biological richness and essential contribution to ecosystem functions. It also offers significant potential for humans, providing valuable ecosystem services such as living resources, energy and climate regulation.
1. Introduction
Key components and structure
From the Latin pelagus, the pelagic domain refers to the open ocean environment, consisting of all the organisms that live within the water column, rather than on the ocean floor or near the coast. A key characteristic is the high mobility and dynamism of its organisms, which are not anchored to substrate and move through the water column. Their movements link habitats and drive processes like benthopelagic coupling (see figure I). An exception to that mobility is some mesopelagic species that are non-migratory, particularly the Cyclothone genus that are largely responsible for what are known as the deep scattering layers, ubiquitous features of the mesopelagic Ref 136 and bathypelagic depths Ref 114.
The pelagic domain is influenced by physical and biogeochemical factors that create distinct and dynamic habitats for marine life at different depths (see figure I). It is subdivided into zones based on light penetration: epipelagic (0-200m), mesopelagic (200-1,000m), bathypelagic (1,000-4,000m), abyssopelagic (4,000-6,000m) and hadalpelagic ( more than 6,000m) zones. This classification aids its description although oversimplification may misrepresent ecosystem processes Ref 39.
Past drivers and pressures
Since the 1800s, the key factors controlling the pelagic domain underwent significant shifts, driven by human-induced climate change. These changes are altering species distributions and productivity, biodiversity and the overall health of the oceans Ref 163, with significant socioecological and economic consequences. Human activities like intense fishing, energy production and global trade are also increasingly affecting the pelagic zone (see figure I), with rising impacts now reaching even remote and once-pristine areas Ref 111.
2. Scientific advances since the first and second World Ocean Assessment
New discoveries and technological advances
Advancements in observational technologies have enhanced the ability to monitor Earth and wildlife, utilizing high-resolution data from sensors in various environments Ref 76 Ref 101. Research vessels now serve as motherships for robots, enabling extensive data collection and multidisciplinary studies Ref 104. The Biogeochemical Argo programme has expanded global insights into ocean productivity and dissolved oxygen Ref 68 Ref 70 Ref 144.
Environmental DNA techniques have enhanced biodiversity understanding across ecosystems Ref 31 Ref 170. They provide new insights into the diets of pelagic species, particularly informing the contribution of gelatinous zooplankton Ref 33. On the other hand, advancements in the study of trophic ecology and ecosystem functioning include compound- specific stable isotope analyses Ref 49 Ref 141.
New assessments and modelling efforts
The latest Earth system models from the Coupled Model Intercomparison Project have projected greater warming, acidification, deoxygenation and nitrate reductions in the ocean, with smaller declines in primary production compared with the previous model Ref 88. The latest models also better capture extreme events like marine heatwaves Ref 120. Species distribution models and marine ecosystem models now integrate complex datasets and environmental dynamics, simulating marine biomass on a global scale Ref 95 Ref 150. These models have revealed uncertainties and help project ecosystem responses to climate changes Ref 19 Ref 59 Ref 151. Recent advancements in functional trait standardization have aided environmental DNA and their applications in ecosystem modelling, connecting ecosystem components based on functional roles and properties Ref 53 Ref 58 Ref 112.
Emergent drivers and effects
Recent analyses have shown how changes in energy transfer efficiency due to ocean warming may lead to reduced biomass of fish and marine mammals, a phenomenon known as trophic amplification Ref 8 Ref 95.
Extreme ocean events have received increased attention due to their abrupt nature and severe impacts. Studies show spatial overlap and temporal synchronicity of compound extreme events, which exacerbate their effects Ref 3. Recent advancements in ocean reanalysis models enable the analysis of vertical structures of these events Ref 165 Ref 169.
Emergent pollutants, transported through oceanic and atmospheric means, also pose an increasing risk to pelagic systems. Global levels may be exceeding planetary boundaries Ref 124.
3. Description of the current state and recent trends
Changes in physical and chemical states
Global ocean temperatures are rising, especially at high latitudes (see figure II) Ref 48 Ref 146 (see sect. 4, chap. 3). In addition, natural variability, such as the Pacific Decadal Oscillation and El Niño Southern Oscillation, affects regional trends Ref 72. The 2023 El Niño led to record sea surface temperatures, with the global average 0.23℃ higher than in 2022, while Antarctic sea ice extent reached unprecedented lows (Copernicus, 2023).
Warming trends also increase the frequency and intensity of marine heatwaves Ref 29 Ref 108 (see figure III). Marine heatwaves affect marine ecosystems by altering species distributions and food web efficiency Ref 140. They can trigger low-oxygen events, worsening ocean deoxygenation Ref 92, leading to habitat compression Ref 77.
The ocean absorbs anthropogenic CO2, leading to increased acidity (decreased pH) and decrease aragonite saturation state Ref 48 Ref 97 (see sect. 4, chap. 3). Ocean salinity and circulation are also changing (see sect. 4, chap. 3) impacting the pelagic domain (see figure II) Ref 21 Ref 34 Ref 35 Ref 45 Ref 54.
Changes in marine productivity and chlorophyll-a concentration
Marine productivity has declined across ocean basins, primarily due to ocean warming, increased stratification and reduced oxygen and nutrient fluxes Ref 22 (see sect. 4, chap. 3). Recent satellite data indicate ongoing declines in chlorophyll-a concentrations, affecting the reproductive performance of marine predators, in particular in the Northern Hemisphere Ref 149. While strong natural variability masks climate-change effects on phytoplankton, trends in ocean colour reflectance are more detectable, revealing that low-latitude oceans have become greener Ref 23. Negative extreme events decrease at high latitudes, while positive extremes intensify, especially along the circumpolar current (see figure IV).
Human activities
Plastic debris is increasingly found in marine organisms Ref 94 Ref 99 Ref 161. Plastics affect marine ecosystems by influencing carbon sinking rates and oxygen levels Ref 87. Humans are exposed to plastics through contaminated seafood Ref 1 Ref 91 Ref 99.
While small-scale trials for mesopelagic fish exist, they are not a significant target for commercial fishing, with total reported catches at 2.7 million tons since the 1950s Ref 113. Economic viability is limited by distance to fishing areas, high fuel costs, rapid fish deterioration and low fish density Ref 84 Ref 110 Ref 157. Rising demand for fishmeal and improved storage methods may make mesopelagic fisheries profitable in the future Ref 110.
Key zones
The mesopelagic zone (200-1,000 m) covers 60% of the surface of the Earth and accounts for 20% of the ocean's volume. This community is vital to the biological carbon pump and the global carbon cycle, transporting carbon to depth through diel vertical migration Ref 4 Ref 81 and mediating the flux of sinking carbon-containing particles Ref 12 (subsect. 5B, chap. 1). Mesopelagic fish and zooplankton contribute 16% each to the global biological carbon pump, corresponding to the sinking particulate organic carbon (POC) flux from surface waters Ref 5 Ref 133. Climate-driven changes are altering mesopelagic communities, impacting the biological carbon pumpRef 81.
Primary production in eastern boundary upwelling systems is disrupted by climate change, as sea surface temperatures rises and upwelling winds intensify. Shifts in pelagic species are observed in upwelling systems, such as the California Current and Southern Benguela Ref 18 Ref 43.
The Arctic and Antarctic regions are highly vulnerable to climate change, in particular in their oceans and cryosphere. Changes in sea ice conditions are leading to profound shifts in the magnitude and phenology of pelagic marine productivity pulses, typical of highly seasonal polar systems, with far-reaching implications for polar ecosystems and their roles in global biogeochemical cycles (see sect. 4, chap. 3 and subchap. 5K) Ref 164.
Key species groups
Climate-driven declines in phytoplankton size and biomass have led to greater losses in higher trophic organisms like fish and marine mammals across the open ocean Ref 60 Ref 95. Ocean warming has also shifted zooplankton distribution, favoured smaller species and altered seasonal patterns of productivity Ref 16 Ref 123. These shifts have large impacts on fish recruitment Ref 13 Ref 69 Ref 74 Ref 117 Ref 143 (see sect. 4, subchap. 4A).
Small pelagic fish contribute over 15% of energy flowing to top predators, supporting 22% of seabirds and 15% of marine mammals in assessed ecosystems Ref 131. Small pelagic fish represent 43% of total fish production and 18% of global catches, Ref 131. Their worldwide status is complex and regionally variable (FAO, 2024). Historically, fluctuations in small pelagic fish populations have been linked to environmental variability, while high fishing pressure and competition among pelagic organisms play crucial roles Ref 2 Ref 28 Ref 64. These fluctuations significantly affect fisheries and predatory species of conservation concern. In 2023, El Niño conditions reduced the habitats of cold-water species like Peruvian anchoveta (Engraulis ringens), the world's largest monospecific fishery, leading to a 50% reduction in industrial landings compared with 2022. Such losses impacted local economies and global aquaculture due to the fishery's role in producing fishmeal and fish oil (see sect. 4, chap. 4).
Deep-pelagic fish are crucial for oceanic carbon sequestration Ref 39 Ref 82 Ref 133 but they face threats from pollutants Ref 42 Ref 79, potential exploitation from fisheries (Hidalgo and Browman, 2019; Pauly and others, 2021), the offshore oil and gas industry Ref 55 and from the emerging practice of deep-sea mining Ref 85.
The open ocean catch was estimated at 11 million tons per year in 2022, primarily consisting of species with open ocean distributions (FAO, 2024). Global tuna catches reached 8.3 million tons in 2022, with 87% of world tuna stocks considered sustainably fished (FAO, 2024). Skipjack tuna (Katsuwonus pelamis) is the third-largest catch among harvested species, while some previously overfished stocks, like bluefin tuna (Thunnus thynnus), have recovered, demonstrating the benefits of effective management Ref 78.Not all species and stocks are assessed, however, in particular small tunas Ref 96.
Chondrichthyans are highly vulnerable to overfishing and bycatch due to slow growth and low reproductive rates Ref 38. Recent trends do not indicate major recoveries Ref 78 and many shark species are threatened with extinction, with three quarters of studied species being at risk Ref 38.
Despite recent recoveries Ref 36, marine mammals in pelagic ecosystems face significant conservation challenges from human activities and climate change (see sect. 4, subchap. 4E). Key threats include bycatch, vessel collisions, plastic and noise pollution and habitat degradation that affects food availability and migratory patterns Ref 10 Ref 44. Many seabird species are also in decline Ref 17, with invasive species, overfishing and climate change as the primary threats Ref 32 Ref 50 Ref 149 (see chap. 4, subchap. 4G). The pelagic phase of marine turtles is largely under-studied Ref 125 and while many populations are declining, a few are increasing Ref 56 (see sect. 4, subchap. 4F).
Effects of changes for humans
Catches from wild-capture fisheries have remained at around 90 million tons per year since the 1980s (FAO, 2024), while commercial fisheries now cover nearly all pelagic ocean areas (Rousseau and others, 2024) (see subsect. 5A, subchaps. 1A and 1B). This expansion may obscure warming-induced shifts in species richness from equatorial waters since the 1970s Ref 26. With global demand for marine protein expected to double by 2050, pressure on pelagic ecosystems will likely increase and move northwards Ref 30.
The blue economy is projected to grow to $3 trillion annually by 2030 Ref 75, while facing challenges due to insufficient ocean knowledge Ref 107. It heavily relies on fisheries, with small pelagic fish and tuna comprising a significant portion of catches (FAO, 2024). Intensive fisheries management has generally improved abundance of several stocks Ref 63. However, increased interest in exploiting deeper waters could jeopardize essential species, leading to declines in biomass and diversity Ref 6 Ref 39 Ref 93.
Pelagic ecosystems suffer from accumulated pollutants, particularly persistent ones like polychlorinated biphenyls and per- and polyfluoroalkyl substances (PFASs), which do not decompose naturally and cause carcinogenesis, teratogenesis and endocrine disruption Ref 166. Nanoplastics and microplastics are of concern, contaminating marine organisms and posing health risks to humans as they bioaccumulate in the food web Ref 41 Ref 134. Risks to the open ocean include light and noise pollution and sediment discharges from mining operations, which contain harmful particulates and chemical additives Ref 25 Ref 37. Mining plumes may impact fisheries Ref 155 and human health through the bioaccumulation of toxins.
Evolution of management and conservation
Managing the high seas is challenging because of their vast and dynamic nature and the complex impact of climate change. Regional fishery management organizations (RFMOs) play crucial roles in managing pelagic species and addressing bycatch, and they are expanding to incorporate ecosystem and climate change considerations (see subsect. 5A, subchap. 1A) Ref 138. Proposing marine protected areas (MPAs) in the pelagic zone faces obstacles due to species' migratory behaviour, impacts of climate change, technological gaps and enforcement difficulties. Recent technological advances have improved monitoring capabilities, although the observation of biological variables and processes still presents important challenges (see sect. 4, chap. 2).
The International Seabed Authority (ISA) is responsible for the regulation of deep-sea exploration and exploitation for minerals in areas beyond national jurisdiction but has yet to adopt a regulatory code for exploitation activities. In 2023, the Authority's Assembly adopted a road map with a view to the adoption during its thirtieth session. Ref 66. The full regulatory framework and time-frame for ISA approval are uncertain Ref 116. The unprecedented geographic scale of deep-sea mining, along with new technologies and poorly understood ecosystems, complicates environmental impact assessments.
The Agreement under the United Nations Convention on the Law of the Sea on the Conservation and Sustainable Use of Marine Biological Diversity of Areas beyond National Jurisdiction aims to protect marine biodiversity in international waters by area-based management tools, including MPAs, and mandate environmental impact assessments. It establishes a framework for sustainable use of ocean resources, enhancing global cooperation to manage the unique challenges of pelagic ecosystems. The Agreement on Marine Biological Diversity of Areas beyond National Jurisdiction was adopted on 19 June 2023 by the Intergovernmental Conference on Marine Biodiversity of Areas Beyond National Jurisdiction convened under the auspices of the United Nations, and ratified on 19 September 2025. The Agreement on Marine Biological Diversity of Areas beyond National Jurisdiction becomes the third implementing agreement to the United Nations Convention on the Law of the Sea, which marked a significant step in ocean conservation and governance.
4. Region-specific issues
Arctic Ocean
The Arctic is experiencing rapid climate transformations, known as Arctic Amplification Ref 24, fuelled by sea ice loss and radiative feedbacks Ref 71. Sea ice extent has significantly declined (Ivanov, 2023) and the Barents Sea has become a "hot spot" for climate change, marked by intensified inflow of warmer, saltier Atlantic waters, known as "Atlantification", which further accelerates sea ice loss Ref 139 (see sect. 4, subchap. 5K). These changes affect biodiversity and abundance of marine communities Ref 57 Ref 58. The region seems to be a sink for contaminants and litter from global sources, transported from lower latitudes Ref 65.
North Atlantic Ocean, Baltic Sea, Black Sea, Mediterranean and North Sea
The North Atlantic faced strong to severe marine heatwaves in the summer of 2023, causing a significant decrease in chlorophyll-a concentration (up to 50 to 60%) in the Eastern North Atlantic, which had a notable impact on ocean productivity. A persistent cold anomaly in the subpolar gyre and low salinity along the Greenland coast, likely due to increased precipitation and ice sheet melting, have altered the distribution of several species (mackerels, bluefin tuna, humpback whales, common eiders, great cormorants) (Josey and others, 2018). In the North Sea, warming surface waters are affecting plankton, fish, and fisheries distribution, with the 0 to 200 m depth zone heavily trawled. While most commercial fish stocks are stable, seabird populations are declining and seal numbers are increasing. In the Baltic Sea, climate-driven changes are reshaping ecosystem. While most pelagic fish stocks are increasing, with the exception of the Baltic herring, non-indigenous species (NIS) are spreading. The Baltic population of the harbour porpoise is critically endangered and presently confined to the western basin, with a dramatic population decline caused by bycatch in fishing gear, underwater noise, chemical pollution and prey depletion. Mediterranean sea surface temperatures have shown positive anomalies Ref 135, with February 2023 reaching a record high of 1.2°C. Rising temperatures and declining primary productivity have affected small pelagic fish, altering their distribution and reducing catches Ref 28 and the spread of non-native pelagic species is changing the biodiversity Ref 168. In the Black Sea, the situation remains poorly assessed, but some large pelagic species, like bluefin tuna and swordfish, were slowly returning before recent extreme events.
South Atlantic Ocean and Wider Caribbean
The Southwestern Atlantic Ocean is highly vulnerable to marine heatwavesRef 7. These marine heatwaves contributed to massive fish mortality and toxic algal blooms in the summer of 2017 Ref 98. Moreover, on particular occasions, marine heatwaves coincided with a severe drought in Southeast Brazil such as in January 2014 Ref 127. In South Africa, management trade-offs between the conservation needs of the endangered African penguin and small pelagic purse seine fisheries have sparked intense debate and legal battles Ref 27 Ref 102.
The increase in Sargassum seaweed blooms (see sect. 4, subchap. 4I) is now stretching across the great Atlantic Sargassum belt from the Gulf of Mexico to the western Atlantic. These blooms are exacerbated by warmer sea temperatures and nutrients runoff from coastal areas and create hypoxic zones affecting marine species, local tourism and fisheries Ref 100 Ref 126.
Indian Ocean, Arabian Sea, Bay of Bengal, Red Sea, Gulf of Aden and Persian Gulf
Warming ocean temperatures and oxygen depletion in the Indian Ocean affect pelagic species, leading to shifts in the distribution of tuna, sardines and mackerel, with species moving towards cooler southern waters Ref 159. For example, in the Arabian Sea, low-oxygen zones are expanding, reducing habitat for pelagic organisms such as Indian mackerel and scads Ref 89. The Bay of Bengal faces increased hypoxia, threatening key pelagic fish species due to changing monsoon patterns Ref 158. In the Persian Gulf, the effects of climate change impacts pelagic fish populations Ref 80.
North Pacific Ocean
This region is increasingly contaminated by microplastics Ref 156 Ref 167. In the western North Pacific, plankton and small pelagic fish biomass fluctuations respond to the Pacific Decadal Oscillation at a multidecadal scale. However, the sea surface temperatures regime has been unconventional in the last decade Ref 86. Despite a negative Pacific Decadal Oscillation phase around 2010, lower sea surface temperatures were recorded in some seasons and small pelagic fish population dynamics have not followed past trends.
South Pacific Ocean
Global warming and rising sea surface temperatures affects marine life, leading to major changes in currents and upwelling zones Ref 21 Ref 147 Ref 160. These changes affect the distribution, reproduction and behavior of many marine animals Ref 9 Ref 105 Ref 115. Concerns about anthropogenic stressors persist, including fisheries Ref 109. The ubiquity of microplastics Ref 11 Ref 20 and the spread of the H5N1 avian influenza virus are affecting seabirds and marine mammals Ref 47 Ref 90 Ref 145.
Southern Ocean
Circumpolar deep-water warms and CO2 concentrations have been rising Ref 106. Atmospheric warming and declining sea-ice extent is most pronounced around the Antarctic Peninsula Ref 50 (see sect. 4, subchap. 5K). This affects the recruitment success of Antarctic krill, a keystone species Ref 74. While current and future physical and biogeochemical changes may increase primary production Ref 118, the food web responses will depend on zooplankton reactions to warming and acidification, affecting higher trophic levels Ref 74 Ref 148.
The long history of resource exploitation, primarily driven by Antarctic krill and toothfish fisheries, remains low compared to global averages Ref 111. However, advancements in technology and favourable environmental conditions could increase the accessibility and pressure on these resources Ref 129. Climate change exacerbates these challenges Ref 50, alongside rising tourism, pollution, pathogens and disturbances from invasive species Ref 14. The growing vulnerability of marine life in the Antarctic has sparked a recent initiative aiming to strengthen humankind's duty in supporting Southern Ocean ecosystems.
5. Key knowledge and capacity gaps
Gaps in key processes
Although there has been progress Ref 39 Ref 40, the mesopelagic zone remains data poor. Increased interest in deep-water ecosystems highlights their ecological significance, alongside significant knowledge gaps and rising threats. Much of the biodiversity below 1,000 m also remains undiscovered, with gaps in taxonomy (smaller organisms being less studied) and species interactions.
Little is known about species movements and ecological interactions among organisms across trophic levels and regions, hindering the understanding of species distribution, ecosystem changes and effective management and conservation effects. Understanding these interactions is crucial for advancing the comprehension of ocean ecosystems and the impact of management actions Ref 137. Technological advancements like in situ imaging and environmental DNA are starting to address some of these important gaps.
The cumulative impact of extreme climate events is becoming a critical research focus, particularly regarding compound events and their influence on ocean dynamics, biodiversity and human livelihoods. Moreover, these events are exacerbated by other anthropogenic impacts on ocean ecosystems, such as pollution and fishing. As the frequency and intensity of extreme events rise, it is essential to understand their effects at various spatiotemporal scales. For instance, the interplay of global warming (centennial scale), Atlantic Multidecadal Oscillation (multidecadal scale), Pacific Decadal Oscillation, and El Niño and La Niña events, along with marine heatwaves (annual scale), could intricately affect organism and community dynamics in ocean ecosystems.
The start of deep-sea mining operations may soon pose a new threat to the deep pelagic communities of the open ocean that play a crucial role as prey for higher trophic levels and in the global biological carbon pump. Assessing the severity of this threat is challenging. Moreover, the interactive effects of multiple stressors on pelagic organisms, populations and communities (and dependencies with benthic counterparts) present a key knowledge gap that requires further research Ref 119. Fisheries impacts and value chains and food nutrition with a pelagic origin, are important knowledge gaps, going beyond the impacts of climate change on fish production Ref 57.
Technological gaps
The pelagic domain remains among the least explored areas on Earth. There is still a critical need for enhanced observational data. Biogeochemical parameters are poorly sampled compared to physical variables. Most monitoring efforts focus on surface-level measurements, limiting understanding of the deep ocean's role in heat and carbon storage, especially below 2,000 m. Recent studies using deep Argo floats reveal that decadal heat content trends are significant in these underexplored regions Ref 73. Gaps remain in measuring vertical ocean velocities, particularly in sub-mesoscale processes. The launch of the Surface Water and Ocean Topography satellite in December 2022 is a significant advancement Ref 46.
Challenges remain in obtaining quantitative estimates of fish and plankton biomass. An estimated 81% of zooplankton monitoring data are either unavailable or only partially accessible Ref 123. There is a pressing need to implement findable, accessible, interoperable and reusable data standards in pelagic data Ref 162 for biodiversity assessments Ref 14 Ref 122. Further efforts on environmental DNA techniques are needed to build reference libraries Ref 123.
Capacity-building needs
Another challenge is about digitizing and mobilizing existing data and knowledge Ref 103 Ref 128. Enhancing the collaboration in international data-sharing programmes through partnerships and capacity-building can improve marine biodiversity monitoring. Taxonomic training, especially for less-known non-vertebrate groups, should remain a priority, as the value of digitized collections depends on accurate species identifications and taxonomic completeness.
Quantitative socioecological assessments are essential for analysing pelagic systems and understanding human impacts. Socioecological models must be scientifically robust yet accessible to managers and decision-makers Ref 61. Despite progress in quantifying marine modelling uncertainty Ref 132 Ref 142, further development is still needed.
Figure I Schematic representation of the pelagic domain

Figure II Temperature trend for the period (a)1982-2017 and (b) 1982-2023; salinity trend for the period (c) 2011-2017 and (d) 2011-2023; chlorophyll-a trend for the period (e) 1998-2017 and (f) 1998-2023
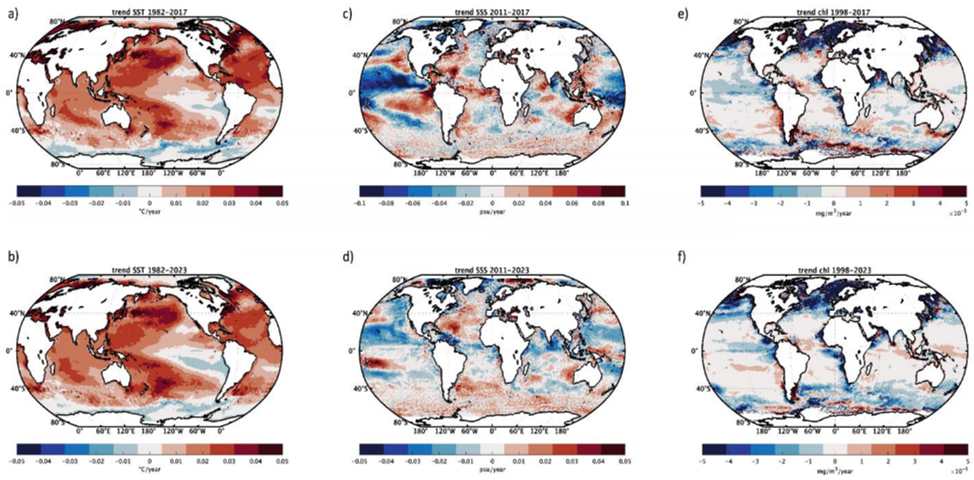
Figure III (a) Mean marine heatwave intensity for the period 2018-2023; (b) same as (a) but for marine heatwave duration; (c) difference between mean marine heatwaves intensity computed over the period 2018-2023 and mean marine heatwave intensity computed over the period 1982-2017; (d) same as (c) but for marine heatwave duration
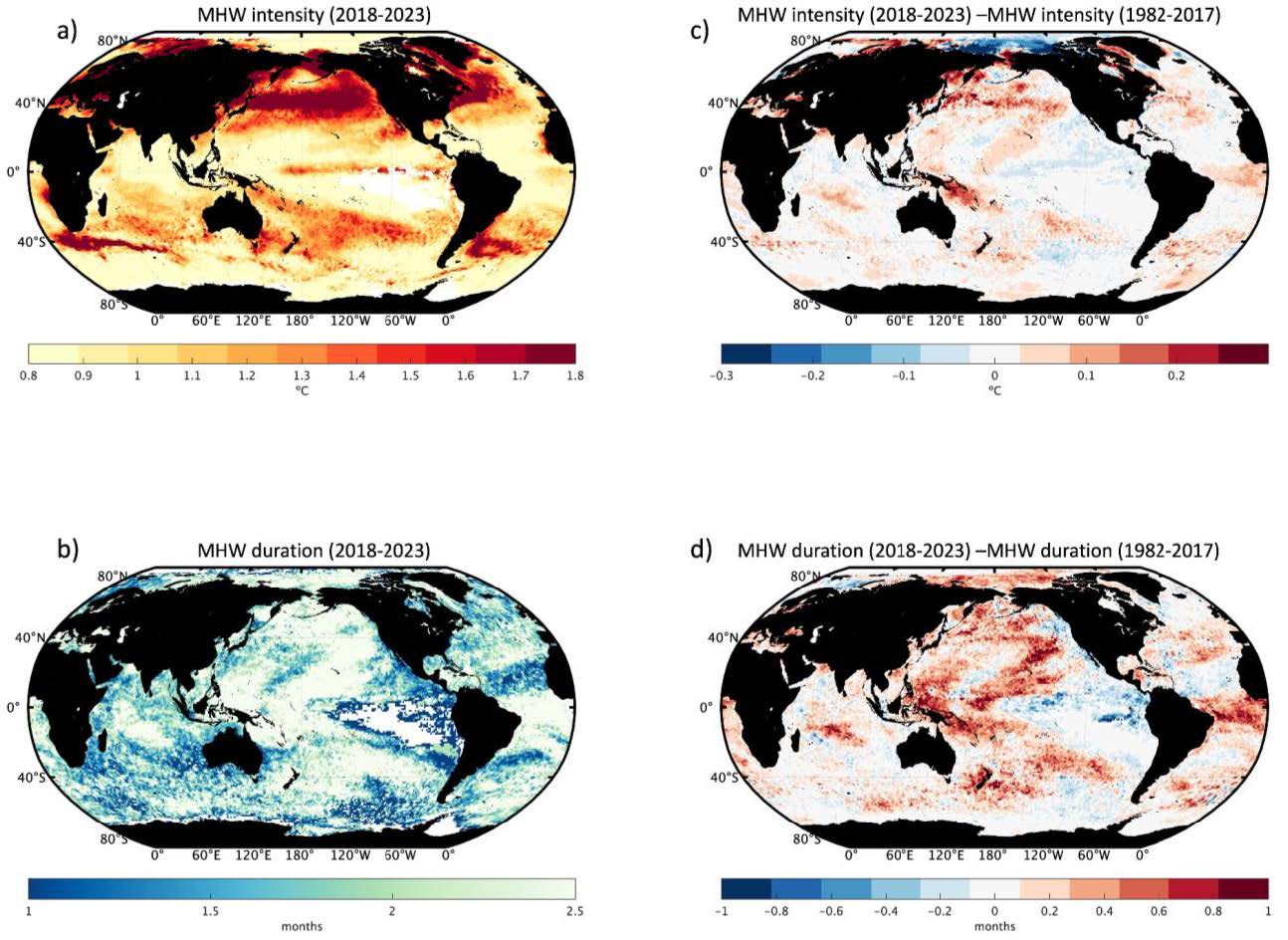
Figure IV (a) Negative and (b) positive extreme events of Chlorophyll-a intensity for the period 2018-2023: (c) differences between negative extreme events of chlorophyll computed over the period 2018-2023 and 2011-2017. Blue colours indicate that negative extreme events were more intense during 2011- 2017. (d) Same as (c) but for positive extreme events of chlorophyll.
References
- Akhbarizadeh, R., Dobaradaran, S., Nabipour, I., Tajbakhsh, S., Darabi, A.H., Spitz, J. (2020). Abundance, composition, and potential intake of microplastics in canned fish. Marine Pollution Bulletin, 160: 111633.
- Alheit, J., Peck, M.A. (2019). Drivers of dynamics of small pelagic fish resources: biology, management and human factors. Marine Ecology Progress Series, 617: 1-6.
- Amaya, D.J., Jacox, M.G., Alexander, M.A., Scott, J.D., Deser, C., Capotondi, A., and others (2023). Bottom marine heatwaves along the continental shelves of North America. Nature Communications, 14: 1038.
- Anderson, T.R., Martin, A.P., Lampitt, R.S., Trueman, C.N., Henson, S.A., Mayor, D.J. (2019). Quantifying carbon fluxes from primary production to mesopelagic fish using a simple food web model. ICES Journal of Marine Science, 76: 690-701.
- Archibald, K.M., Siegel, D.A., Doney, S.C. (2019). Modeling the impact of zooplankton diel vertical migration on the carbon export flux of the biological pump. Global Biogeochemical Cycles, 33: 181-199.
- Ariza, A., Lengaigne, M., Menkes, C., Lebourges-Dhaussy, A., Receveur, A., Gorgues, T., and others (2022). Global decline of pelagic fauna in a warmer ocean. Nature Climate Change, 12: 928-934.
- Artana, C., Rodrigues, R.R., Fevrier, J., Coll, M. (2024). Characteristics and drivers of marine heatwaves in the western South Atlantic. Communications Earth & Environment, 5.
- Atkinson, A., Rossberg, A.G., Gaedke, U., Sprules, G., Heneghan, R.F., Batziakas, S., and others (2024). Steeper size spectra with decreasing phytoplankton biomass indicate strong trophic amplification and future fish declines. Nature Communications, 15: 381.
- Avila, I.C., Dormann, C.F., García, C., Payán, L.F., Zorrilla, M.X. (2020). Humpback whales extend their stay in a breeding ground in the Tropical Eastern Pacific. ICES Journal of Marine Science,; 77: 109-118.
- Avila, I.C., Kaschner, K., Dormann, C.F. (2018). Current global risks to marine mammals: taking stock of the threats. Biological Conservation, 221: 44-58.
- Bakir, A., Desender, M., Wilkinson, T., Van Hoytema, N., Amos, R., Airahui, S., and others (2020). Occurrence and abundance of meso and microplastics in sediment, surface waters, and marine biota from the South Pacific region. Marine Pollution Bulletin, 160: 111572.
- Baumas, C.M., Le Moigne, F.A., Garel, M., Bhairy, N., Guasco, S., Riou, V., and others (2021). Mesopelagic microbial carbon production correlates with diversity across different marine particle fractions. The ISME Journal, 15: 1695-1708.
- Beaugrand, G., Brander, K.M., Alistair Lindley, J., Souissi, S., Reid, P.C. (2003). Plankton effect on cod recruitment in the North Sea. Nature, 426: 661-664.
- Becker, S.L., Boyd, C., Handley, J.M., Raymond, B., Reisinger, R., Ropert-Coudert, Y., and others (2024). Scaling up ocean conservation through recognition of key biodiversity areas in the Southern Ocean from multispecies tracking data. Conservation Biology, e14345.
- Bednaršek, N., Feely, R.A., Pelletier, G., Desmet, F. (2023). Global synthesis of the status and trends of ocean acidification impacts on shelled pteropods. Oceanography, 36: 130-137.
- Benedetti, F., Vogt, M., Elizondo, U.H., Righetti, D., Zimmermann, N.E., Gruber, N. (2021). Major restructuring of marine plankton assemblages under global warming. Nature communications, 12: 5226.
- BirdLife International (2018). State of the world's birds: taking the pulse of the planet. BirdLife International.
- Blamey, L.K., Shannon, L.J., Bolton, J.J., Crawford, R.J., Dufois, F., Evers-King, H., and others (2015). Ecosystem change in the southern Benguela and the underlying processes. Journal of Marine Systems, 144: 9-29.
- Blanchard, J.L., Novaglio, C. (2024). Climate change risks to marine ecosystems and fisheries - Projections to 2100 from the Fisheries and Marine Ecosystem Model Intercomparison Project. In Blanchard, J.L., Novaglio, C., eds. FAO Fisheries and Aquaculture Technical Paper. 707. FAO, Rome.
- Bleszynski, M., Clark, E. (2023). Preliminary Assessment into the Prevalence and Distribution of Microplastics in North and South Pacific Island Beaches. Microplastics, 2: 219-229.
- Bograd, S.J., Jacox, M.G., Hazen, E.L., Lovecchio, E., Montes, I., Pozo Buil, M., and others (2023). Climate change impacts on eastern boundary upwelling systems. Annual Review of Marine Science, 15: 303-328.
- Boyce, D.G., Lewis, M.R., Worm, B. (2010). Global phytoplankton decline over the past century. Nature, 466: 591-596.
- Cael, B., Bisson, K., Boss, E., Dutkiewicz, S., Henson, S. (2023). Global climate-change trends detected in indicators of ocean ecology. Nature, 619: 551-554.
- Carmack, E., Polyakov, I., Padman, L., Fer, I., Hunke, E., Hutchings, J., and others (2015). Toward quantifying the increasing role of oceanic heat in sea ice loss in the new Arctic. Bulletin of the American Meteorological Society, 96: 2079-2105.
- Christiansen, S., Bräger, S. (2023). Developing best environmental practice for polymetallic nodule mining-a review of scientific recommendations. Frontiers in Marine Science, 10: 1243252.
- Chust, G., González Taboada, F., Fernandes-Salvador, J.A., Cheung, W., Coll, M. (2024). Climate change impacts on marine fish ecology and fisheries. In Cabral H, ed. Ecology of Marine Fishes. Elsevier, pp. 29.
- Cochrane, K., Butterworth, D., Hilborn, R., Parma, A., Plagányi, É., Sissenwine, M. (2024). Errors and bias in marine conservation and fisheries literature: Their impact on policies and perceptions. Marine Policy, 168: 106329.
- Coll, M., Bellido, J.M., Pennino, M.G., Albo-Puigserver, M., Báez, J.C., Christensen, V., and others (2024). Retrospective analysis of the pelagic ecosystem of the Western Mediterranean Sea: drivers, changes and effects. Science of The Total Environment, 167790.
- Collins, M., Sutherland, M., Bouwer, L., Cheong, S., Frölicher, T., Jacot Des Combes, H., and others (2022). Extremes, abrupt changes and managing risks. IPCC special report on the ocean and cryosphere in a changing climate. Cambridge University Press, pp. 589-655.
- Cruz, L., Pennino, M., Lopes, P. (2024). Fisheries track the future redistribution of marine species. Nature Climate Change, 1-8.
- Deeg, C.M., Li, S., Esenkulova, S., Hunt, B.P., Schulze, A.D., Miller, K.M. (2023). Environmental DNA survey of the Winter Salmonosphere in the Gulf of Alaska. Environmental DNA, 5: 519-539.
- Dias, M.P., Martin, R., Pearmain, E.J., Burfield, I.J., Small, C., Phillips, R.A., and others (2019). Threats to seabirds: a global assessment. Biological Conservation, 237: 525-537.
- Dischereit, A., Throm, J.K., Werner, K.M., Neuhaus, S., Havermans, C. (2024). A belly full of jelly? DNA metabarcoding shows evidence for gelatinous zooplankton predation by several fish species in Greenland waters. Royal Society Open Science, 11: 240797.
- Ditlevsen, P., Ditlevsen, S. (2023). Warning of a forthcoming collapse of the Atlantic meridional overturning circulation, Nat. Commun., 14, 4254.
- Douville, H., Cheng, L. (2024). Asymmetric sea surface salinity response to global warming:“Fresh gets fresher but salty hesitates". Geophysical Research Letters, 51: e2023GL107944.
- Duarte, C.M., Agusti, S., Barbier, E., Britten, G.L., Castilla, J.C., Gattuso, J-P., and others (2020). Rebuilding marine life. Nature, 580: 39-51.
- Duarte, C.M., Chapuis, L., Collin, S.P., Costa, D.P., Devassy, R.P., Eguiluz, V.M., and others (2021). The soundscape of the Anthropocene ocean. Science, 371: eaba4658.
- Dulvy, N.K., Pacoureau, N., Rigby, C.L., Pollom, R.A., Jabado, R.W., Ebert, D.A., and others (2021). Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Current Biology, 31: 4773-4787. e8.
- Eduardo, L.N., Mincarone, M.M., Sutton, T., Bertrand, A. (2024). Deep-Pelagic Fishes Are Anything But Similar: A Global Synthesis. Ecology Letters, 27: e14510.
- Egorova, Y., Reygondeau, G., Cheung, W.W., Pakhomov, E.A. (2024). Species Distribution Models for Mesopelagic Mesozooplankton Community. Journal of Biogeography.
- Fauvelle, V., Garel, M., Tamburini, C., Nerini, D., Castro-Jiménez, J., Schmidt, N., and others (2021). Organic additive release from plastic to seawater is lower under deep-sea conditions. Nature Communications, 12: 4426.
- Ferreira, G.V., Justino, A.K., Eduardo, L.N., Schmidt, N., Martins, J.R., Ménard, F., and others (2023). Influencing factors for microplastic intake in abundant deep-sea lanternfishes (Myctophidae). Science of The Total Environment, 867: 161478.
- Fiechter, J., Pozo Buil, M., Jacox, M.G., Alexander, M.A., Rose, K.A. (2021). Projected shifts in 21 st century sardine distribution and catch in the California Current. Frontiers in Marine Science, 8: 685241.
- Fortuna, C., Fortibuoni, T., Bueno-Pardo, J., Coll, M., Franco, A., Giménez, G., and others (2024). Top predator status and trends: ecological implications, monitoring and mitigation strategies to promote ecosystem-based management. Frontiers in Marine Science - Marine Ecosystem Ecology.
- Fox, M.D., Guillaume-Castel, R., Edwards, C.B., Glanz, J., Gove, J.M., Green, J.M., and others (2023). Ocean currents magnify upwelling and deliver nutritional subsidies to reef-building corals during El Niño heatwaves. Science Advances, 9: eadd5032.
- Fu, L.L., Pavelsky, T., Cretaux, J.F., Morrow, R., Farrar, J.T., Vaze, P., and others (2024). The surface water and ocean topography mission: A breakthrough in radar remote sensing of the ocean and land surface water. Geophysical Research Letters, 51: e2023GL107652.
- Gamarra-Toledo, V., Plaza, P.I., Gutiérrez, R., Inga-Diaz, G., Saravia-Guevara, P., Pereyra-Meza, O., and others (2023). Mass mortality of sea lions caused by highly pathogenic avian influenza A (H5N1) virus. Emerging Infectious Diseases, 29: 2553.
- Garcia-Soto, C., Cheng, L., Caesar, L., Schmidtko, S., Jewett, E.B., Cheripka, A., and others (2021). An overview of ocean climate change indicators: Sea surface temperature, ocean heat content, ocean pH, dissolved oxygen concentration, arctic sea ice extent, thickness and volume, sea level and strength of the AMOC (Atlantic Meridional Overturning Circulation). Frontiers in Marine Science, 8: 642372.
- Gimenez, J., Albo-Puigserver, M., Laiz-Carrión, R., Lloret-Lloret, E., Bellido, J.M., Coll, M. (2023). Trophic position variability of European sardine by compound-specific stable isotope analyses. Canadian Journal of Fisheries and Aquatic Sciences, 80. https://doi.org/10.1139/cjfas-2022-019.
- Gimeno, M., Giménez, J., Chiaradia, A., Lloyd, D., Seddon, P., Ropert-Coudert, Y., and others (2024). Climate and human stressors on global penguin hotspots: current assessments for future conservation. Global Change Biology, 30: e17143.
- Gordó-Vilaseca, C., Costello, M.J., Coll, M., Jüterbock, A., Reiss, H., Stephenson, F. (2024). Future trends of marine fish biomass distributions from the North Sea to the Barents Sea. Nature Communications, 15: 5637.
- Gordó-Vilaseca, C., Stephenson, F., Coll, M., Lavin, C., Costello, M.J. (2023). Three decades of increasing fish biodiversity across the north-east Atlantic and the Arctic Ocean. Proceedings of the National Academy of Science of the United States of America, 120: e2120869120.
- Green, S.J., Brookson, C.B., Hardy, N.A., Crowder, L.B. (2022). Trait-based approaches to global change ecology: moving from description to prediction. Proceedings of the Royal Society B, 289: 20220071.
- Gutierrez-Guerra, M.Á., Pérez-Hernández, M.D., Vélez-Belchí, P. (2024). Intensified upwelling: normalized sea surface temperature trends expose climate change in coastal areas. Ocean Science, 20: 1291-1308.
- Hawkins, S., Todd, P., Russell, B., Lemasson, A., Allcock, A., Byrne, M., and others (2023). Review of the central and south atlantic shelf and deep-sea benthos: science, policy, and management. Oceanography and Marine Biology: An annual review, volume 61, 61: 127-218.
- Hays, G.C., Schofield, G., Papazekou, M., Chatzimentor, A., Katsanevakis, S., Mazaris, A.D. (2024). A pulse check for trends in sea turtle numbers across the globe. Iscience, 27.
- Heneghan, R.F,, Everett, J.D., Blanchard, J.L., Sykes, P., Richardson, A.J. (2023). Climate-driven zooplankton shifts cause large-scale declines in food quality for fish. Nature Climate Change, 13: 470- 477.
- Heneghan, R.F., Everett, J.D., Sykes, P., Batten, S.D., Edwards, M., Takahashi, K., and others (2020). A functional size-spectrum model of the global marine ecosystem that resolves zooplankton composition. Ecological Modelling, 435: 109265.
- Heneghan, R.F., Galbraith, E.D., Blanchard, J.L., Harrison, C., Barrier, N., Bulman, C., and others (2021). Disentangling diverse responses to climate change among global marine ecosystem models. Progress in Oceanography, 198.
- Henson, S.A., Laufkötter, C., Leung, S., Giering, S.L., Palevsky, H.I., Cavan, E.L. (2022). Uncertain response of ocean biological carbon export in a changing world. Nature Geoscience, 15: 248-254.
- Heymans, J.J., Bundy, A., Christensen, V., Coll, M., De Mutsert, K., Fulton, E., and others (2020). The Ocean Decade: A true ecosystem modelling challenge. Frontiers in Marine Science - Marine Fisheries, Aquaculture and Living Resources. 10.3389/fmars.2020.554573.
- Hidalgo, M., Browman, H.I. (2019). Developing the knowledge base needed to sustainably manage mesopelagic resources. 76. Oxford University Press, pp. 609-615.
- Hilborn, R., Amoroso, R.O., Anderson, C.M., Baum, J.K., Branch, T.A., Costello, C., and others (2020). Effective fisheries management instrumental in improving fish stock status. Proceedings of the National Academy of Sciences.
- Hilborn, R., Buratti, C.C., Díaz Acuña, E., Hively, D., Kolding, J., Kurota, H., and others (2022). Recent trends in abundance and fishing pressure of agency-assessed small pelagic fish stocks. Fish and Fisheries, 23: 1313-1331.
- ICES (2022). Central Arctic Ocean ecoregion - Ecosystem Overview. Report of the ICES Advisory Committee. ICES Advice 2022, Section 2.1. https://doi.org/10.17895/ices.advice.21731240.
- ISA (2024). ISA Assembly concludes its twenty-ninth session. International Seabed Authority Press Release, 28 June. www.isa.org.jm/news/.
- Ivanov V. (2023). Arctic sea ice loss enhances the oceanic contribution to climate change. Atmosphere, 14: 409.
- Izett, R.W., Fennel, K., Stoer, A.C., Nicholson, D.P. (2024). Reviews and syntheses: expanding the global coverage of gross primary production and net community production measurements using Biogeochemical-Argo floats. Biogeosciences, 21: 13-47.
- Jaspers, C., Hopcroft, R.R., Kiørboe, T., Lombard, F., López-Urrutia, Á., Everett, J.D., and others (2023). Gelatinous larvacean zooplankton can enhance trophic transfer and carbon sequestration. Trends in ecology & evolution, 38: 980-993.
- Jemai, A., Wollschläger, J., Voß, D., Zielinski, O. (2021). Radiometry on Argo floats: From the multispectral state-of-the-art on the step to hyperspectral technology. Frontiers in Marine Science, 8: 676537.
- Jenkins, M., Dai, A. (2021). The impact of Sea-Ice loss on Arctic climate feedbacks and their role for Arctic Amplification. Geophysical Research Letters, 48: e2021GL094599.
- Johnson, G.C., Lyman, J.M. (2020). Warming trends increasingly dominate global ocean. Nature Climate Change 2020, 10: 757-761.
- Johnson, G.C., Purkey, S.G. (2024). Refined estimates of global ocean deep and abyssal decadal warming trends. Geophysical Research Letters, 51: e2024GL111229.
- Johnston, N.M., Murphy, E.J., Atkinson, A., Constable, A.J., Cotté, C., Cox, M., and others (2022). Status, change, and futures of zooplankton in the Southern Ocean. Frontiers in Ecology and Evolution, 9: 624692.
- Jolly, C. (2016). The ocean economy in 2030. Proceedings of the Workshop on Maritime Cluster and Global Challenges 50th Anniversary of the WP6, Paris, France. 1.
- Joo, R., Picardi, S., Boone, M.E., Clay, T.A., Patrick, S.C., Romero-Romero, V.S., and others (2022). Recent trends in movement ecology of animals and human mobility. Movement ecology, 10: 26.
- Jorda, G., Marbà, N., Bennett, S., Santana-Garcon, J., Agusti, S., Duarte, C.M. (2020). Ocean warming compresses the three-dimensional habitat of marine life. Nature Ecology & Evolution, 4: 109-114.
- Juan-Jordá, M.J., Murua, H., Arrizabalaga, H., Merino, G., Pacoureau, N., Dulvy, N.K. (2022). Seventy years of tunas, billfishes, and sharks as sentinels of global ocean health. Science, 378: eabj0211.
- Justino, A.K., Ferreira, G.V., Schmidt, N., Eduardo, L.N., Fauvelle, V., Lenoble, V., and others (2022). The role of mesopelagic fishes as microplastics vectors across the deep-sea layers from the Southwestern Tropical Atlantic. Environmental Pollution, 300: 118988.
- Khalfeh Nilsaz, M., Aliabadi, S., Savari, A., Dehghan Mediseh, S. (2024). Predict the effects of climate change on primary production and vulnerability of fisheries species in coastal waters of the northern Persian Gulf. Iranian Journal of Fisheries Sciences, 23: 893-909.
- Kiko, R., Bianchi, D., Grenz, C., Hauss, H., Iversen, M., Kumar, S., and others (2020). Zooplankton and nekton: Gatekeepers of the biological pump. 7. Frontiers Media SA, pp. 545.
- Klevjer, T.A., Irigoien, X., Røstad, A., Fraile-Nuez, E., Benítez-Barrios, V.M., Kaartvedt, S. (2016). Large scale patterns in vertical distribution and behaviour of mesopelagic scattering layers. Scientific reports, 6: 19873.
- Kolodziejczyk, N., Hamon, M., Boutin, J., Vergely, J., Reverdin, G., Supply, A., Reul, N. (2021) Objective analysis of SMOS and SMAP sea surface salinity to reduce large-scale and time-dependent biases from low to high latitudes. Journal of Atmospheric and Oceanic Technology, vol. 38, issue 3. 405-421.
- Kourantidou, M., Jin, D. (2022). Mesopelagic-epipelagic fish nexus in viability and feasibility of commercial-scale mesopelagic fisheries. Natural Resource Modeling, 35: e12350.
- Kung, A., Svobodova, K., Lèbre, E., Valenta, R., Kemp, D., Owen, J.R. (2021). Governing deep sea mining in the face of uncertainty. Journal of Environmental Management, 279: 111593.
- Kuroda, H., Saito, T., Kaga, T., Takasuka, A., Kamimura, Y., Furuichi, S., and others (2020). Unconventional sea surface temperature regime around Japan in the 2000s-2010s: potential influences on major fisheries resources. Frontiers in Marine Science, 7: 574904.
- Kvale, K., Hunt, C., James, A., Koeve, W. (2023). Regionally disparate ecological responses to microplastic slowing of faecal pellets yields coherent carbon cycle response. Frontiers in Marine Science, 10: 1111838.
- Kwiatkowski, L., Torres, O., Bopp, L., Aumont, O., Chamberlain, M., Christian, J.R., and others (2020). Twenty-first century ocean warming, acidification, deoxygenation, and upper-ocean nutrient and primary production decline from CMIP6 model projections. Biogeosciences, 17: 3439-3470.
- Lachkar, Z., Levy, M., Hailegeorgis, D., Vallivattathillam, P. (2023). Differences in recent and future trends in the Arabian Sea oxygen minimum zone: processes and uncertainties. Frontiers in Marine Science, 10: 1122043.
- Leguia, M., Garcia-Glaessner, A., Muñoz-Saavedra, B., Juarez, D., Barrera, P., Calvo-Mac, C., and others (2023). Highly pathogenic avian influenza A (H5N1) in marine mammals and seabirds in Peru. Nature Communications, 14: 5489.
- Leslie, H.A., Van Velzen, M.J., Brandsma, S.H., Vethaak, A.D., Garcia-Vallejo, J.J., Lamoree, M.H. (2022). Discovery and quantification of plastic particle pollution in human blood. Environment international, 163: 107199.
- Li C, Huang J, Liu X, Ding L, He Y, Xie Y. (2024). The ocean losing its breath under the heatwaves. Nature Communications, 15: 6840.
- Lin, C-H., Wei, C-L., Ho, S.L., Lo, L. (2023). Ocean temperature drove changes in the mesopelagic fish community at the edge of the Pacific Warm Pool over the past 460,000 years. Science Advances, 9: eadf0656.
- Liu, Y., Ayaka, M., Toshikazu, K., Helena, F., Takashi, M. (2023). Analysis of microplastics in the intestines of stranded cetaceans. AACL Bioflux, 16: 2746-2756.
- Lotze, H.K., Tittensor, D.P., Bryndum-Buchholz, A., Eddy, T.D., Cheung, W.L., Galbraith ED, and others (2019). Global ensemble projections reveal trophic amplification of ocean biomass declines with climate change. Proceedings of the National Academy of Science of the United States of America, 116: 12907-12912.
- Lucena-Frédou, F., Mourato, B., Frédou, T., Lino, P.G., Muñoz-Lechuga, R., Palma, C., and others (2021). Review of the life history, fisheries, and stock assessment for small tunas in the Atlantic Ocean. Reviews in Fish Biology and Fisheries, 31: 709-736.
- Ma, D., Gregor, L., Gruber, N. (2023). Four decades of trends and drivers of global surface ocean acidification. Global Biogeochemical Cycles, 37: e2023GB007765.
- Manta, G., de Mello, S., Trinchin, R., Badagian, J., Barreiro, M. (2018). The 2017 record marine heatwave in the southwestern Atlantic shelf. Geophysical Research Letters, 45: 12,449-12,456.
- Marmara, D., Katsanevakis, S., Brundo, M-V., Tiralongo, F., Ignoto, S., Krasakopoulou, E. (2023). Microplastics ingestion by marine fauna with a particular focus on commercial species: a systematic review. Frontiers in Marine Science, 10: 1240969.
- Marsh, R., Skliris, N., Tompkins, E.L., Dash, J., Dominguez, Almela, V., Tonon, T., and others (2023). Climate-sargassum interactions across scales in the tropical Atlantic. PLOS Climate, 2: e0000253.
- McCauley, D.J., Andrzejaczek, S., Block, B.A., Cavanaugh, K.C., Cubaynes, H.C., Hazen, E.L., and others (2024). Improving Ocean Management Using Insights from Space. Annual Review of Marine Science, 17.
- McInnes, AM, Weideman EA, Carpenter-Kling T, Barham P, Christian M, Day K, and others Commercial fishery no-take zones for African penguins minimize fisheries losses at the expense of conservation gains. ICES Journal of Marine Science 2024; 81: 1632-1646.
- Meyer, C., Kreft, H., Guralnick, R., Jetz, W., Boitani, L. (2015). Global priorities for an effective information basis of biodiversity distributions. Nat Commun, 6: 8221.
- Miller, A., Virmani, J.I. (2023). Advanced marine technologies for ocean research. Deep Sea Research Part II: Topical Studies in Oceanography, 105340.
- Montalva, F., Pavés, H., Pérez-Venegas, D., Barrientos-E., K.G., Valencia, C., Miranda-Urbina, D., and others (2022). Lower marine productivity increases agonistic interactions between sea lions and fur seals in Northern Pacific Patagonia. Current Zoology, 68: 657-666.
- Morley, S.A., Abele, D., Barnes, D.K., Cárdenas, C.A., Cotté, C., Gutt, J., and others (2020). Global drivers on southern ocean ecosystems: changing physical environments and anthropogenic pressures in an earth system. Frontiers in Marine Science, 7: 547188.
- Narwal, S., Kaur, M., Yadav, D.S., Bast, F. (2024). Sustainable blue economy: Opportunities and challenges. Journal of Biosciences, 49: 18.
- Oliver, E.C., Benthuysen, J.A., Darmaraki, S., Donat, M.G., Hobday, A.J., Holbrook, N.J., and others (2021). Marine heatwaves. Annual review of marine science, 13: 313-342.
- Ortiz, M., Hermosillo-Núñez, B. (2024). Quantifying stability and resilience of eco-social keystone species complexes for coastal marine ecosystems of the Caribbean Sea and eastern Pacific: applications in conservation and monitoring programmes. Philosophical Transactions B, 379: 20230176.
- Paoletti S, Nielsen JR, Sparrevohn CR, Bastardie F, Vastenhoud BM. Potential for mesopelagic fishery compared to economy and fisheries dynamics in current large scale Danish pelagic fishery. Frontiers in Marine Science 2021; 8: 720897.
- Paolo, F.S., Kroodsma, D., Raynor, J., Hochberg, T., Davis, P., Cleary, J., and others (2024). Satellite mapping reveals extensive industrial activity at sea. Nature, 625: 85-91.
- Pata, P.R., Hunt, B.P. (2023). Harmonizing marine zooplankton trait data toward a mechanistic understanding of ecosystem functioning. Limnology and Oceanography.
- Pauly, D., Piroddi, C., Hood, L., Bailly, N., Chu, E., Lam, V., and others (2021). The biology of mesopelagic fishes and their catches (1950-2018) by commercial and experimental fisheries. Journal of Marine Science and Engineering, 9: 1057.
- Peña, M., Munuera-Fernández, I., Nogueira, E., González-Quirós, R. (2021). First recording of a bathypelagic deep scattering layer in the Bay of Biscay. Progress in oceanography, 198: 102669.
- Peters, K.J., Stockin, K.A., Saltré, F. (2022). On the rise: Climate change in New Zealand will cause sperm and blue whales to seek higher latitudes. Ecological Indicators, 142: 109235.
- Pickens, C., Lily, H., Harrould-Kolieb, E., Blanchard, C., Chakraborty, A. (2024). From what-if to what- now: Status of the deep-sea mining regulations and underlying drivers for outstanding issues. Marine Policy, 169: 105967.
- Pinchuk, A.I., Batten, S.D., Strasburger, W.W. (2021). Doliolid (Tunicata, Thaliacea) blooms in the southeastern Gulf of Alaska as a result of the recent marine heat wave of 2014-2016. Frontiers in Marine Science, 8: 625486.
- Pinkerton, M.H., Boyd, P.W., Deppeler, S., Hayward, A., Höfer, J., Moreau, S. (2021). Evidence for the impact of climate change on primary producers in the Southern Ocean. Frontiers in Ecology and Evolution, 9: 592027.
- Pinsky, M.L., Selden, R.L., Kitchel, Z.J. (2020). Climate-driven shifts in marine species ranges: scaling from organisms to communities. Annual review of marine science, 12: 153-179.
- Qiu, Z., Qiao, F., Jang, C.J., Zhang, L., Song, Z. (2021). Evaluation and projection of global marine heatwaves based on CMIP6 models. Deep Sea Research Part II: Topical Studies in Oceanography, 194: 104998.
- Rahmstorf S. (2024). Is the Atlantic Overturning Circulation approaching a tipping point? Oceanography, 37: 16-29.
- Ramírez, F., Sbragaglia, V., Soacha, K., Coll, M., Piera, J. (2022). Challenges for Marine Ecological Assessments: Completeness of Findable, Accessible, Interoperable, and Reusable Biodiversity Data in European Seas. Frontiers in Marine Science, section Ocean Observation.
- Ratnarajah, L., Abu-Alhaija, R., Atkinson, A., Batten, S., Bax, N.J., Bernard, K.S., and others (2023). Monitoring and modelling marine zooplankton in a changing climate. Nature Communications, 14: 564.
- Richardson, K., Steffen, W., Lucht, W., Bendtsen, J., Cornell, S.E., Donges, J.F., and others (2023). Earth beyond six of nine planetary boundaries. Science advances, 9: eadh2458.
- Robinson, N.J., Aguzzi, J., Arias, S., Gatto, C., Mills, S.K., Monte, A., and others (2023). Global trends in sea turtle research and conservation: Using symposium abstracts to assess past biases and future opportunities. Global Ecology and Conservation, 47: e02587.
- Robledo, D., Vázquez-Delfín, E., Freile-Pelegrín, Y., Vásquez-Elizondo, R.M., Qui-Minet, Z.N., Salazar- Garibay, A. (2021). Challenges and opportunities in relation to Sargassum events along the Caribbean Sea. Frontiers in Marine Science, 699664.
- Rodrigues, R.R., Taschetto, A.S., Sen Gupta, A., Foltz, G.R. (2019). Common cause for severe droughts in South America and marine heatwaves in the South Atlantic. Nature Geoscience, 12: 620-626.
- Rogers, A.D., Appeltans, W., Assis, J., Ballance, L.T., Cury, P., Duarte, C., and others (2022). Discovering marine biodiversity in the 21st century. Advances in Marine Biology, 93: 23-115.
- Rogers, A.D., Frinault, B.A., Barnes, D., Bindoff, N.L., Downie, R., Ducklow, H.W., and others (2020). Antarctic futures: an assessment of climate-driven changes in ecosystem structure, function, and service provisioning in the Southern Ocean. Annual Review of Marine Science, 12: 87-120.
- Rousseau, Y., Blanchard, J.L., Novaglio, C., Pinnell, K.A., Tittensor, D.P., Watson, R.A., and others (2017). A database of mapped global fishing activity 1950-2017. Scientific Data, 11: 48.
- Ruzicka, J., Chiaverano, L., Coll, M., Garrido, S., Tam, J., Murase, H., and others (2024). The role of small pelagic fish in diverse ecosystems: knowledge gleaned from food-web models. Marine Ecology Progress Series, SPF2av9.
- Rynne, N., Novaglio, C., Blanchard, J.L., Bianchi, D., Christensen, V., Coll, M., and others (2025). A Skill Assessment Framework for the Fisheries and Marine Ecosystem Model Intercomparison Project. Earth's Future, 13.
- Saba, G.K., Burd, A.B., Dunne, J.P., Hernández-León, S., Martin, A.H., Rose, K.A., and others (2021). Toward a better understanding of fish-based contribution to ocean carbon flux. Limnology and Oceanography, 66: 1639-1664.
- Sala, B., Gimenez, J., Fernandez-Arribas, J., Bravo, C., Lloret Lloret, E., Esteban, A., and others (2022). Organophosphate ester plasticizers in edible fish from the Mediterranean Sea: Marine pollution and human exposure. Environmental Pollution, 292.
- Salat, J., Pascual, J., Flexas, M., Chin, T.M., Vazquez-Cuervo, J. (2019). Forty-five years of oceanographic and meteorological observations at a coastal station in the NW Mediterranean: a ground truth for satellite observations. Ocean Dynamics, 69: 1067-1084.
- Sarmiento-Lezcano, A.N., Olivar, M.P., Caballero, M.J., Couret, M., Hernandez-Leon, S., Castellón, A., and others (2023). Swimbladder properties of Cyclothone spp. in the northeast Atlantic Ocean and the Western Mediterranean Sea. Frontiers in Marine Science, 10: 1093982.
- Schaber, M., Gastauer, S., Cisewski, B., Hielscher, N., Janke, M., Peña, M., and others (2022). Extensive oceanic mesopelagic habitat use of a migratory continental shark species. Scientific Reports, 12: 2047.
- SCRS-ICCAT (2024). Report of the standing committee on research and statistics (SCRS). ICCAT 2024. https://www.iccat.int/Documents/Meetings/Docs/2024/Reports/2024_SCRS_ENG.pdf.
- Skagseth, Ø., Eldevik, T., Årthun, M., Asbjørnsen, H., Lien, V.S., Smedsrud, L.H. (2020). Reduced efficiency of the Barents Sea cooling machine. Nature Climate Change 2020; 10: 661-666.
- Smith, K.E., Burrows, M.T., Hobday, A.J., King, N.G., Moore, P.J., Sen Gupta, A., and others (2023). Biological impacts of marine heatwaves. Annual Review of Marine Science, 15: 119-145.
- Stahl, A.R., Rynearson, T.A., McMahon, K.W. (2023). Amino acid carbon isotope fingerprints are unique among eukaryotic microalgal taxonomic groups. Limnology and Oceanography, 68: 1331-1345.
- Steenbeek, J., Ortega, P., Bernardello, R., Christensen, V., Coll, M., Exarchou, E., and others (2024). Making ecosystem modelling operational - a novel distributed execution framework to systematically explore ecological responses to divergent climate trajectories. Earth's Future, 12: e2023EF004295.
- Steinberg, D.K., Landry, M.R. (2017). Zooplankton and the ocean carbon cycle. Annual review of marine science, 9: 413-444.
- Stoer, A.C., Takeshita, Y., Maurer, T.L., Begouen Demeaux, C., Bittig, H.C., Boss, E., and others (2023). A census of quality-controlled Biogeochemical-Argo float measurements. Frontiers in Marine Science, 10: 1233289.
- Stokstad, E. (2023). Deadly avian flu hits Galapagos Concerns rise for boobies, finches, and other endemic species. Science, 381: 1382.
- Storto, A., Yang, C. (2024). Acceleration of the ocean warming from 1961 to 2022 unveiled by large- ensemble reanalyses. Nature Communications, 15: 545.
- Sutton, P., Rickard, G., Roemmich, D. (2024). Southwest Pacific Ocean warming driven by circulation changes. Geophysical Research Letters, 51: e2024GL109174.
- Swadling, K.M., Constable, A.J., Fraser, A.D., Massom, R.A., Borup, M.D., Ghigliotti, L., and others (2023). Biological responses to change in Antarctic sea ice habitats. Frontiers in Ecology and Evolution, 10: 1073823.
- Sydeman, W.J., Schoeman, D.S., Thompson, S.A., Hoover, B.A., García-Reyes, M., Daunt, F., and others (2021). Hemispheric asymmetry in ocean change and the productivity of ecosystem sentinels. Science, 372: 980-983.
- Tittensor, D.P., Eddy, T.D., Lotze, H.K., Galbraith, E.D., Cheung, W., Barange, M., and others (2018). A protocol for the intercomparison of marine fishery and ecosystem models: Fish-MIP v1.0 Geoscientific Model Development, 11: 1421-1442.
- Tittensor, D.P., Novaglio, C., Harrison, C.S., Heneghan, R.F., Barrier, N., Bianchi, D., and others (2021). Next-generation Earth System Models reveal higher climate risks for marine ecosystems. Nature Climate Change, 11: 973-981.
- TMC (2025). TMC Announces June 27, 2025 Submission Date for Subsidiary NORI's ISA Application, and Expanded Company Strategy. Nov 12, 2024, Globe Newswire, New York.
- https://investors.metals.co/news-releases/news-release-details/tmc-announces-june-27-2025-submission- date-subsidiary-noris-isa.
- Van de Waal, D.B., Litchman, E. (2020). Multiple global change stressor effects on phytoplankton nutrient acquisition in a future ocean. Philosophical Transactions of the Royal Society B, 375: 20190706.
- Van Der Grient, J., Drazen, J. (2021). Potential spatial intersection between high-seas fisheries and deep- sea mining in international waters. Marine Policy, 129: 104564.
- Van Sebille, E., Wilcox, C., Lebreton, L., Maximenko, N., Hardesty, B.D., Van Franeker, J.A., and others (2015). A global inventory of small floating plastic debris. Environmental Research Letters, 10: 124006.
- Vastenhoud, B.M., Bastardie, F., Andersen, K.H., Speirs, D.C., Nielsen, J.R. (2023). Economic viability of a large vessel mesopelagic fishery under ecological uncertainty. Frontiers in Marine Science, 10: 1285793.
- Vidhya, V., Jyothibabu, R., Jagadeesan, L., Rashid, C., Alok, K., Arunpandi, N., and others (2022). Oxygen minimum zone copepods in the Arabian Sea and the Bay of Bengal: Their adaptations and status. Progress in Oceanography, 206: 102839.
- Waller, M.J., Humphries, N.E., Womersley, F.C., Loveridge, A., Jeffries, A.L., Watanabe, Y., and others (2024). The vulnerability of sharks, skates, and rays to ocean deoxygenation: Physiological mechanisms, behavioral responses, and ecological impacts. Journal of Fish Biology.
- Wang, S., Jing, Z., Wu, L., Sun, S., Peng, Q., Wang, H., and others (2023). Southern hemisphere eastern boundary upwelling systems emerging as future marine heatwave hotspots under greenhouse warming. Nature Communications, 14: 28.
- Wieczorek, A.M., Morrison, L., Croot, P.L., Allcock, A.L., MacLoughlin, E., Savard, O., and others (2018). Frequency of microplastics in mesopelagic fishes from the Northwest Atlantic. Frontiers in Marine Science, 5: 339138.
- Wilkinson, M.D., Dumontier, M., Aalbersberg, I.J., Appleton, G., Axton, M., Baak, A., and others (2016). The FAIR Guiding Principles for scientific data management and stewardship. Scientific data, 3: 1-9.
- Williamson, P., Guinder, V.A. (2021). Effect of climate change on marine ecosystems. The impacts of climate change, 115-176.
- Willis, M.D., Lannuzel, D., Else, B., Angot, H., Campbell, K., Crabeck, O., and others (2023). Polar oceans and sea ice in a changing climate. Elementa, 11.
- Wong, J., Münnich, M., Gruber, N. (2024). Column-compound extremes in the global ocean. AGU Advances, 5: e2023AV001059.
- Worm, B., Lotze, H.K., Jubinville, I., Wilcox, C., Jambeck, J. (2017). Plastic as a persistent marine pollutant. Annual Review of Environment and Resources, 42.
- Yu, R-S., Singh, S. (2023). Microplastic pollution: threats and impacts on global marine ecosystems. Sustainability, 15: 13252.
- Zenetos, A., Tsiamis, K., Galanidi, M., Carvalho, N., Bartilotti, C., Canning-Clode, J., and others (2022). Status and trends in the rate of introduction of marine non-indigenous species in European seas. Diversity, 14: 1077.
- Zhang, Y., Du, Y., Feng, M., Hobday, A.J. (2023). Vertical structures of marine heatwaves. Nature Communications, 14: 6483.
- Zhang, Y., Pavlovska, M., Stoica, E., Prekrasna, I., Yang, J., Slobodnik, J., and others (2020). Holistic pelagic biodiversity monitoring of the Black Sea via eDNA metabarcoding approach: From bacteria to marine mammals. Environment International, 135: 105307.