Fjord systems
Writing team: Jose L. Iriarte (coordinating author), Ian Butler (co-lead member), Agneta Fransson, Jennifer M. Jackson, Lorenz Meire, Renato Quinones (lead member), Sofia Ribeiro and Fiammetta Straneo.
Key points
- Fjords are ecologically carbon organic-rich regions and home to Indigenous Peoples and local communities.
- Global warming is already affecting both northern and southern hemisphere fjords (through warming, deoxygenation and acidification).
- The loss of glacier ice and the changing seasonality of the hydrological cycle is affecting physical and chemical factors and processes in fjord systems.
1. Introduction
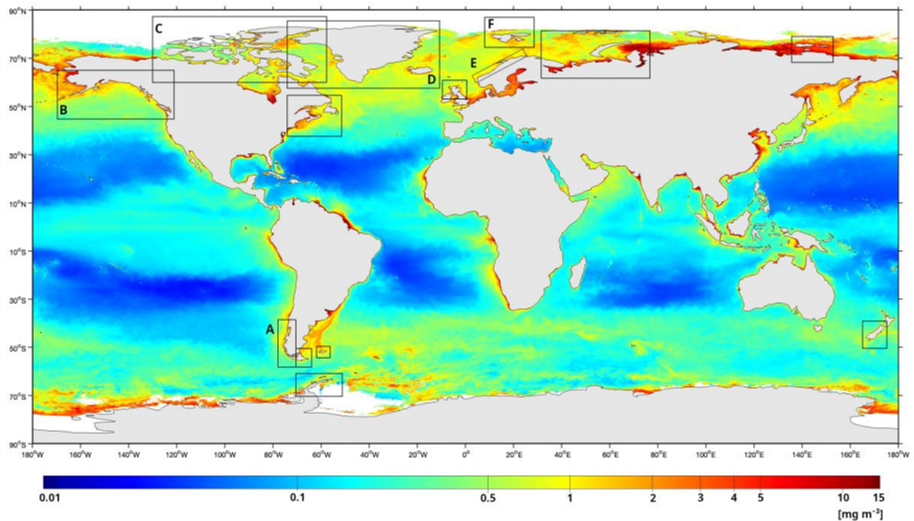
The fjord systems of the world are characterized by a deep basin connecting directly or indirectly with the open sea at the mouth Ref 85. Fjords are often long, narrow, deep and steep-sided inlets, frequently branched and sinuous, or straight, and may contain one or more submarine sills (a shallow dome of bedrock that forms at the mouth of a fjord). The presence and size of these sills at the fjord's mouth can restrict the exchange of water between the fjord and the coastal ocean, often leading to highly stratified conditions that result in low-oxygen bottom waters. Main fjord regions occur along the coasts of North and South America (above 42° latitude); many sub-Antarctic islands; the southwest coast of New Zealand's South Island; Antarctica; the Russian and Canadian Arctic Archipelagos; Svalbard, Norway; Iceland; Greenland; and northern Europe, including the west coast of Scotland Ref 97 Ref 116 Ref 19 (see figure I). Fjord systems are large transitional coastal zones, encompassing many distinct climate-hydrological-oceanographic interconnected systems, that are strongly influenced by interactions with mountain ranges, freshwater input, local to hemispheric-scale atmospheric weather patterns, glacier ice fields and the open ocean (i.e. Pacific, Atlantic) Ref 11 Ref 62. These transitional coastal zones are characterized by a complex geography including the presence of several marine habitats such as bays, estuaries, gulfs, islands, fjords and channels. Inner coastal waters are influenced by freshwater flux from rivers, which are fed from precipitation, as well as snow, glacier and sea ice melting. The interaction of this freshwater with oceanic waters can generate strong horizontal and vertical salinity, density and nutrient gradients (see sect. 4, chap. 3). Fjords are also carbon cycle hotspots where the largest amount of organic carbon per unit area is buried in sediments, making them key ecosystems for carbon cycle regulation over time Ref 119 Ref 32 Ref 23 Ref 112 Ref 133 Ref 10. At present, coastal fjord regions are increasingly experiencing anthropogenic impacts due to global processes (e.g. climate change) and regional activities (e.g. eutrophication) in inshore waters. Under the framework established in the third World Ocean Assessment, the present subchapter focuses for the first time on several fjord systems of the world (see figure I), with particular emphasis on recent studies in mainland Norway, Svalbard, Greenland, Patagonia and Pacific North America.
Figure I Map showing the global distribution of fjord systems overlaid with map showing chlorophyll-a concentrations. Fjord systems of the world, with particular emphasis on recent studies in Patagonia (A), Pacific North America (B), Canadian Arctic Archipelago (C), Greenland margins (D), mainland Norway (E) and Svalbard (F).
2. Descriptions of the overall status
Mainland Norway
Mainland Norway has thousands of fjords along the coast (58°-72ºN). Fjords on the west coast are influenced by two distinct water masses: the low-salinity Norwegian Coastal Water in the upper layer and the high-salinity Atlantic Water in the lower layer beneath. The vertical structure and horizontal extent of coastal water show seasonal oscillations due to prevailing wind conditions, where alongshore northerly (southerly) winds can cause coastal upwelling (downwelling) Ref 117 Ref 111. Norwegian fjords are affected by land-fjord-shelf interactions such as river water input carrying organic matter and nutrients to the fjords and further to the shelf and some glaciated fjords have meltwater runoff. Some fjords have a shallow sill that prevents frequent renewal of the deep water due to the low salinity (low density) surface layer. Strong stratification and limited vertical mixing can result in long residence times of deep-water masses, which can lead to a decrease in oxygen levels and create challenging environments for local ecosystems Ref 4 Ref 24 Ref 71.
Fjords are productive due to nutrient-rich freshwater runoff from rivers and land promoting primary and secondary production. Diatom-spring blooms are most common in Norwegian fjords and as food for zooplankton. Periodically, spring blooms of the calcium carbonate forming coccolithophore have been reported in several Norwegian fjords Ref 75, observed from space (NASA; Modis). Large copepods are an important food source for fish (e.g. Atlantic coastal cod) and shrimps Ref 132. Some fjords have unique cold-water corals Ref 105. Fixation of CO2 by phytoplankton is a significant factor controlling the partial pressure of CO2 (pCO2), with the development of spring blooms in northernmost fjords. Decreased CO2 uptake is observed in autumn due to a combined reduction of the mixed layer with the entrainment of high-CO2 subsurface water, reduced biological production and increased surface water temperatures Ref 1. Time-series data reveal that the North Atlantic Oscillation significantly influences river discharge, temperature and precipitation in central Norway, making terrigenous input and erosional processes in the fjord hinterland highly sensitive to atmospheric variability. In addition, marine carbonate productivity is the primary source of calcite and calcium in all three fjords Ref 30.
Svalbard (Spitsbergen)
Most fjords on the west coast of Svalbard exhibit a typical fjord circulation associated with a three-layer structure: a fresh surface layer in summer; a layer of intermediate water, transformed Atlantic Water in the middle to outer part; and local fjord water in the deeper part of the fjord Ref 56 Ref 20 Ref 88 Ref 118. The summer surface layer is mainly formed by freshwater from glacial meltwater and river runoff Ref 115. East coast fjords are mostly impacted by cold Arctic water and thicker sea ice than are found in the west coast fjords. Due to the sill depth, some fjords with a shallow sill depth such as Billefjorden have limited inflow of water from outside the fjord to renew the bottom water and creates arctic conditions in the inner parts Ref 125 Ref 123.
Mountains surrounding the fjords have different bedrock compositions where weathering supplies minerals and ions to the fjord Ref 26, which impacts the marine chemistry (alkalinity) and carbonate system (ocean CO2 uptake) Ref 34 Ref 35 Ref 28. Glacier runoff contributes to surface stratification and nutrient supply. Sub-glacial melting affects the presence and supply of nutrients and other bio-essential substances (iron) derived from the bedrock/sediment at the bottom and transported by vertical mixing Ref 60. High primary production blooms in Kongsfjorden (usually in April-June) are initiated due to shallow surface stratification and are often dominated by diatoms (e.g. Ref 51) and when the surface layer becomes more mixed and warmer the blooms are dominated by other species such as Phaeocystis pouchetii (e.g. Ref 58). Submarine melt in Arctic/Svalbard fjords is typically driven by the presence of warm water such as Atlantic Water and its derivative coastal water masses Ref 115 Ref 122. This melting causes low-density water to rise to the surface in the inner fjord, with positive responses on phytoplankton growth (diatoms), zooplankton (large copepods), fish, birds and sea mammals Ref 125 Ref 81 Ref 48 Ref 7. On the other hand, sediments carried by glacial meltwater can lead light availability, thereby limiting primary production Ref 60 Ref 27. Sea ice is important for ice algae (mainly diatoms) which play an important role as an early food source for many marine invertebrates such as polychaetae and roundworms (Nematoda) that are known to reside within sea ice Ref 125. Glacial surface melt is largest in summer and limited during winter, while subglacial freshwater generated all year round through geothermal and frictional heat and delayed release of subglacially-stored freshwater Ref 131. Recent research in Kongsfjorden, Svalbard, reveals how freshwater runoff from marine- and land-terminating glaciers affects the biogeochemical cycles of carbon, nutrients and trace elements in the Arctic fjord system. The data show that intense carbonate weathering in proglacial catchments adds dissolved carbonates to fjord waters, mitigating the reduced buffering capacity from glacial runoff Ref 113. Also observed in other west-Spitsbergen fjords such as Tempelfjorden Ref 34 Ref 36. Work in Smeerenburgfjorden, Kongsfjorden and Van Keulenfjorden, of Western Svalbard, revealed that benthic recycling of glacially derived dissolved iron into seawater, followed by partial re-oxidation and deposition, facilitates iron transport across the fjords and potentially into adjacent shelf waters. This new data on biogeochemical processes and carbon cycling in a climate-sensitive, high-latitude fjord region, aids in predictions of future Arctic ecosystem changes Ref 134.
Greenland glacial fjords
Kalaallit Nunaat (Greenland) is characterized by hundreds of glacial fjords, distributed around its perimeter, that connect the ocean with the ice sheet. Many of these fjords are associated with marine-terminating glaciers that discharge icebergs that move through the fjords and into the coastal ocean. Meltwater released into the fjords from glacier melt, iceberg melt and tundra runoff constitute a significant freshwater export into the coastal ocean Ref 8. One key feature of Greenland's glacial fjords is the release of large volumes of summer surface melt at the grounding zone of marine terminating glaciers, hundreds of meters below sea-level Ref 91 Ref 9. The large density difference between this freshwater and the ocean water gives rise to turbulent, rising plumes, which play a key role in driving the fjord circulation, increase melting of the glacier face and upwell deep nutrients Ref 120 Ref 89 Ref 59 Ref 16. Estimating the dissolved silica (dSi) flux from the Greenland Ice Sheet has been difficult, with existing estimates differing by an order of magnitude. A new study indicates a modest dissolve Si flux from runoff, with smaller contributions from melting icebergs and suspended particles Ref 61. The confluence of Arctic and Atlantic- origin waters flow along-side with the Arctic waters inshore and the Atlantic waters on the continental slope Ref 122. Greenland fjords are characterized by seasonal sea-ice coverage varying significantly with latitude and between east and west Greenland. Sea ice plays a vital role for Indigenous Peoples and local communities, supporting traditional livelihoods such as fishing and hunting Ref 76.
Large phytoplankton blooms occur in spring as solar radiation increases and sea-ice retreats Ref 130. The mixing of different water types and nutrient upwelling associated with glacial plumes contributes to a continued high productivity in the fjords with marine-terminating glaciers during summer Ref 90 Ref 73. These high primary production blooms, often dominated by diatoms, provide a valuable food source for attracting large zooplankton communities and consequently for lower trophic level forage fish species such as capelin Ref 47 Ref 77. These rich marine ecosystems are not only of ecological importance but also of socio- economic importance for commercial fisheries Ref 13 Ref 89 as well as local fishing and hunting activities Ref 107 Ref 121.
Patagonian fjords
Along the Chilean Patagonia (41 - 54°S), the coast is characterized by a complex geography including the presence of fjords and channels. Northern Patagonia (41 - 47ºS) showed high freshwater input by large rivers (up to 600 m3 s-1) along the coast characterized by waters of low salinity and high silicic acid concentration Ref 69 Ref 22. Such discharges affect the nutrient content and their stoichiometric ratio (N:P:Si), as well as the water column structure showing a strong haline stratification Ref 79. Their interaction with the Subantarctic Water (SAAW), generate strong horizontal and vertical salinity (and density) gradients Ref 106 that contribute to maintain the phytoplankton community under high nutrient availability within the euphotic zone Ref 63. The interplay between river discharges and oceanic water masses, as the principal nutrient sources, leads the maximum peak of chlorophyll-a to occur in the inner coastal waters during the austral spring-summer months Ref 64. The pronounced salinity gradient by the riverine freshwater input can be extended to the offshore region, reaching as far as approximately 80°W, generating a strong relationship between stratification, nutrient content and surface total chlorophyll a biomass Ref 38. In contrast, southern Patagonia (47 - 54°S) is strongly affected by western winds (which have maximum intensity during summer) and the fluvial influence is lower compared to snow and glacial melting processes Ref 39 Ref 3. These conditions promote a deeper mixed layer, low silicic acid and nitrate concentrations (Cuevas and others, 2014) and high content of suspended sediments, which affect the light availability in the upper layer, mainly in the inner waters Ref 45 Ref 69. The biogeochemistry of Patagonia's fjords is shaped by complex, changing factors, including fjord geomorphology, local climate, interactions of multiple water masses and atmospheric modes Ref 21 Ref 80 Ref 101 Ref 108. Despite this complexity, studies of water columns, surface sediments and downcore organic carbon emphasize the heterogeneity of carbon inputs and fate Ref 46 Ref 104 Ref 109 Ref 110.
Pacific North American fjords
In North America, fjords are located on all three ocean coasts - the Atlantic, Arctic and Pacific Oceans but here the focus is on Pacific North American fjords, which span Washington State (United States of America), British Columbia (Canada) and Alaska (United States of America). These fjords are part of the Northeast Pacific Coastal Temperate Rainforest Ecoregion, which includes prehumid and seasonal rainforests Ref 12 Ref 62. The Pacific North American fjords are fringed by mountain ranges: the Olympic range in western Washington and the coast mountain range (also known as the Cascade Range) that runs throughout Washington, British Columbia and Alaska. Freshwater flows from the coastal mountains to the coast via thousands of small coastal rivers and five large continental rivers Ref 44. Rivers are observed at the heads of most fjords and the seasonality of the river depends on whether the source of the freshwater is rain, snow or glacial melt Ref 92. In British Columbia, there are 42 fjords that are longer than 18.5 km and of these, the outer sill depth ranges from 6 to 490 m, the maximum depth ranges from 75 to 730 m, and the length ranges from 33 to 130 km Ref 97. In most cases there are three water types in fjords: deep water (greater than sill depth), that in most fjords is seasonally modified by deep water renewal Ref 29; intermediate water (above sill and below surface water), that can be modified by wind-driven or advective processes Ref 67; and surface water that is driven by estuarine circulation Ref 97. In recent research it has been pointed out that the ecosystem within fjords is different than surrounding waters, with high phytoplankton concentrations Ref 84 that are dominated year-round by diatoms Ref 70 Ref 25 and very abundant and diverse zooplankton populations Ref 95.
3. Region-specific changes
Mainland Norway
Freshwater input is increasing to the coastal zone which is an important area for fisheries (cod, salmon, shrimp) and aquaculture. Cod egg dispersion models show negative impacts of increased low-density water with low oxygen Ref 114. There is recent evidence that coastal waters in mainland Norway are becoming lighter due to climate change, resulting in a stronger stratification in the fjord basins with reduced mixing and consequent deoxygenation in fjords Ref 71. An estimated increase of 0.6℃ in temperature in the fjord basins after 1990 showed that basin water renewal occurred less frequently in most fjord basins along the Norwegian coast Ref 71. Masfjorden in southern Norway is one of the fjords showing recent oxygen-depleted basin water Ref 99. Impacts of ocean acidification: some glaciated fjords have meltwater runoff that dilutes buffering ions (and alkalinity), increases CO2-uptake and increases ocean acidification Ref 1 Ref 72. Human activities such as shipping, aquaculture (see subsect. 5A, subchap. 1D), fishing and pollution (e.g. mining waste; see sect. 4, chap. 6) and climate stressors such as ocean warming and acidification, negatively impacting the fjords. Because of the unique and diverse environment, several such fjords (i.e. Ytre Hardangerfjorden fjord) have recently become marine protected areas (MPAs).
Svalbard
Ocean warming of 0.7º C per year in winter and 0.6° C per year in summer is observed on the shelf of western Spitsbergen and Isfjorden Ref 118 and a decline in sea ice is reported in Kongsfjorden Ref 96 Ref 42. Glaciers in Svalbard are melting and retreating and freshwater input to the coastal zone is increasing Ref 74 Ref 43 due to warming (e.g. Ref 33). Fjords in Svalbard experience increased ocean acidification due to increased glacial discharge and meltwater-induced CO2 uptake Ref 15. These waters are further modified by accumulated dissolved inorganic carbon from sea-ice brine rejection and respiration of organic matter which further contributes to acidification Ref 28 Ref 34 Ref 35. Studies in Kongsfjorden (west Spitsbergen) showed inter-annual variability in biogeochemical properties with lower aragonite saturation (Ωar) and pH (indicators of ocean acidification) in cold years with extensive sea-ice cover, relative to warm years with a greater influence from warm and saline Atlantic Water inflow Ref 35. Differences in phytoplankton spring bloom composition in Kongsfjorden were observed in contrasting years where in a cold year with more sea ice the spring bloom was dominated by diatoms and in a warm year, with less sea ice, the spring bloom was dominated by haptophyte Ref 124 Ref 7. In years with less surface stratification, warmer water and deeper mixed layer the blooms were dominated by species such as Phaeocystis pouchetii Ref 58. Moreover, the reported decreased sea ice presence in Svalbard fjords Ref 96 will lead to less ice algae and earlier phytoplankton blooms Ref 125, resulting in changes in the diversity of sea-ice protist (e.g. diatoms) and ice-associated fauna Ref 57. In sediments of Kongsfjorden in Svalbard benthic remineralization could increase nutrient sources like iron (Fe). Interannual variations in primary productivity, expected to intensify with global warming and changes in glacial sediment delivery will affect benthic iron and sulfur cycling. Over time, this may lead to decreased benthic Fe fluxes. With glacial retreat and shifts in productivity, fjords like Kongsfjorden may become less efficient carbon sinks, burying less organic matter in deep sediments Ref 54. Seepage of methane is reported in most Svalbard fjords and natural gas below the permafrost, with methane flares measured in Isfjorden, Kongsfjorden and VanMijenfjorden Ref 55. With warming, permafrost thawing and methane release into the fjords will likely increase with impacts on carbon cycling in the fjords and shelves. Knowledge gaps on the methane dynamics in Svalbard still exist Ref 55.
Greenland's margins
Climate change, including the rapid warming of the Arctic, is resulting in significant changes over Greenland and along its coastal margins. These changes include the retreat of glaciers, increased surface melt Ref 82, reduced sea ice Ref 18 and warming air temperatures Ref 14. These trends are superimposed with long-term modes of variability, such as the Arctic and North Atlantic Oscillations and the Atlantic Multidecadal Oscillation, that account for a significant fraction of the variability in this region Ref 49. Records of physical and chemical properties and of ecosystem and fisheries variability, in the fjords and in the coastal region are, however, scarce - making it challenging to quantify change that has already occurred. Proxy records derived from fjord sediment archives offer valuable insights into past changes beyond the instrumental era Ref 53. Fjord sediments simultaneously record changes in sea ice, ice-sheet discharge, ice-ocean interactions and associated ecosystem responses Ref 102 Ref 128. Proxy-data studies from Greenland fjords from both West and East Greenland have revealed multi-decadal changes in the inflow of Atlantic-derived warm water and its relationship with glacier dynamics Ref 6 Ref 128. They also have revealed that present-day impacts of freshwater discharge on fjord productivity are unprecedented in centuries to millennia Ref 93 Ref 94. The increase in the upwelling of nutrients associated with subglacial discharge may be partly offset by the retreat of glaciers into shallower water - thus reducing the ability to upwell deep nutrients Ref 89 Ref 60. Sea-ice decrease is impacting wintertime travel and fishing, which Indigenous Peoples and local communities rely on for sustenance and threatens the future viability of key polynya ecosystems Ref 103. Sea-ice cover serves as winter refuges for marine mammal species that are harvested by Indigenous People in Greenland fjords Ref 52. At the same time, reduced sea-ice resulted in increased primary productivity as light limitations are reduced and wind-induced mixing increased (e.g. Baffin Bay and the Arctic Ocean) Ref 136 Ref 37.
Patagonia
The Patagonia region is undergoing an increasing anthropogenic affectation due to global phenomena, including hydrological anomalies, glacier retreatment, climatological shifts Ref 3 Ref 2 Ref 39 and regional productive activities such as large-scale aquaculture farms (salmon and mussels) allocated along inshore waters Ref 87. In northern Patagonia (41 - 44°S), climate change projections forecast 20% less rainfall in the next decade Ref 2, which affect negatively the annual dynamics of large river streamflows, water chemistry (silicic acid) and pelagic (mollusk larvae) and benthic organisms inhabiting fjords systems Ref 127 as well as the physical support (weakening of haline stratification and increase of sea surface temperature; Ref 79). Recurrent harmful microorganisms' events observed in coastal and oceanic regions off northern Patagonia since 2016, coincided with the synergy of remote climatic phenomena (El Niño) and global warming that led to very dry conditions, higher than normal solar radiation, high sea surface temperatures, and reduced wind velocities Ref 78 (Mardones and others, 2023). Significant mass loss (65%) in the Patagonia icefields (47 - 55°S) may continue in the future Ref 86 Ref 3, where ocean-terminating and land-terminating fjords are likely exposed to increasing freshwater runoff (freshening effect) and increasing rain precipitation Ref 40. Patagonian fjords and inner seas are affected by human-induced global changes, including climate-ocean events, altered freshwater inputs, harmful algal blooms, settlement, aquaculture, glacial melt and coastal hypoxia (Dussaillant and others, 2019) Ref 46 Ref 65 Ref 100 Ref 80.
Pacific North America
Data of temperature, salinity and oxygen have been collected since 1951 in British Columbia fjords. Recent work found that deep water warmed by 1.2 to 1.4℃ and lost 0.4 to 0.7 mL L-1 of oxygen from 1951 to 2020 Ref 68. These changes are explained by the upwelling of offshore water that was warmed by the 2014-2016 marine heatwave to fjords via deep water renewal Ref 66 Ref 5; the 2014-2016 marine heat wave water lingered in deep fjord water until 2020 Ref 68. Waters in fjords are often acidic Ref 50 and impacted by random events Ref 5.
Glaciers in British Columbia are melting rapidly and it is predicted that the volume of glacial ice will shrink by approximately 70% by 2100 relative to 2005 Ref 17. In addition to altering the amount of freshwater transported to the ocean, as the glaciers melt, they cause extreme events such as the 2020 outburst flood that destroyed salmon habitat and advected tons of inorganic and organic sediments into Bute Inlet Ref 41. There is evidence that these changes to the seasonality and amount of freshwater that enters fjords are impacting the timing of the spring bloom Ref 135 Ref 62 Ref 25, as well as zooplankton dynamics Ref 126.
4. Outlook
General awareness of fjords and the key ecosystem services offered by them are essential to determine actions on management under natural disturbances as well as present and projected climate change scenarios. Linking physical-ecological and socioeconomic structures and their ecosystems functions are key to propose and incorporate both mitigation and adaptation processes such as nature-based solutions and climate refugees. Long-term monitoring approaches designed and coordinated by scientists and stakeholders must be undertaken in existing and future marine and terrestrial protected areas located in fjord systems. In the present subchapter as well as others, such as cold-water corals and sponges (CWCS) (sect. 4, subchap. 5E), estuaries and deltas (sect. 4, subchap. 5F) and small-scale aquaculture (subsect. 5A, subchap. 1D), it has been pointed out that fjord regions are under threat from intensive human activities (including land-use, aquaculture, dams, tourism) and the effects of climate change (including warming, freshening, ice loss, permafrost thawing, erosion and sea level rise).
5. Remaining key knowledge and capacity changes and any new gaps
- A critical knowledge gap exists for fjords habitats, as only a small percentage of the world's fjords have been studied.
- Change from marine-terminating glaciers to land-terminating glaciers would determine changes in surface and sub-glacial melt, sediment supply that limits light, stronger stratification and nutrient limitation for primary production, changes in species composition and, ultimately, the dynamic of carbon flux.
- The contribution to the global carbon cycle of methane emissions from melting ice sheets and glaciers in the Arctic unknown.
- The resilience of fjord systems to changes in hydrologic (precipitation, melting, runoff) and oceanographic (water masses, stratification, nutrient stoichiometry) to present conditions remain a major gap.
- Advances in modelling research, such as in relation to the impacts of freshening, coastal acidification, sea surface warming, and deoxygenation processes, are lacking and need to be compared with in situ studies approaches.
References
- Aalto, N.J., and others (2021). Drivers of Atmosphere-Ocean CO2 Flux in Northern Norwegian Fjords. Frontiers Marine Science. doi: 10.3389/fmars.2021.69209.
- Aguayo, Rodrigo, and others (2019). The glass half-empty: Climate Change drives lower freshwater input in the coastal system of the Chilean northern Patagonia. Climatic Change. https://doi.org/10.1007/s10584- 019-02495-6.
- Aguayo, Rodrigo, and others (2024). Unravelling the sources of uncertainty in glacier runoff projections in the Patagonian Andes (40 - 56S). The Cryosphere, vol. 18, 5383-5406. doi.org/10.5194/tc-18-5383- 2024.
- Aksnes, D.L., and others (2019). Multi-decadal warming of Atlantic water and associated decline of dissolved oxygen in a deep fjord. Estuarine, Coastal and Shelf Science, 228. https://doi.org/10.1016/j.ecss.2019.106392.
- Alin, S.R., and others (2024). Seasonality and response of ocean acidification and hypoxia to major environmental anomalies in the southern Salish Sea, North America (2014-2018). Biogeosciences, vol. 21, 1639-1673. https://doi.org/10.5194/bg-21-1639-2024.
- Andresen, C., and others (2012). Rapid response of Helheim Glacier in Greenland to climate variability over the past century. Nature Geoscience, vol. 5, 37-41.
- Assmy Philipp, and others (2023). Seasonal plankton dynamics in Kongsfjorden during two years of contrasting environmental conditions. Progress in Oceanography, vol. 213, 102996. https://doi.org/10.1016/j.pocean.2023.102996.
- Bamber, J.L., and others (2018). Land ice freshwater budget of the Arctic and North Atlantic Oceans: 1. Data, methods, and results. Journal of Geophysical Research: Oceans, vol. 123, 1827-1837.
- Beaird, N.L., and others (2018). Export of strongly diluted Greenland meltwater from a major glacial fjord. Geophysical Research Letters, vol. 45, 4163-4170. doi: 10.1029/2018GL077000.
- Bertrand, S. (2025). Biogeochemistry of riverine organic matter inputs to the Patagonian fjords and implications for fjord organic carbon budgets. Journal of Geophysical Research, Biogeosciences, vol. 130.
- Bianchi, Thomas, S., and others (2020). Fjords as aquatic critical zones (ACzs). Earth-Science Reviews, vol. 203. https://doi.org/10.1016/j.earscirev.2020.103145.
- Bidlack, A.L., and others (2021). Climate-Mediated Changes to Linked Terrestrial and Marine Ecosystems across the Northeast Pacific Coastal Temperate Rainforest Margin. BioScience, vol. 71, 581- 595. https://doi.org/10.1093/biosci/biaa171.
- Boje, J., and others (2014). Seasonal migration, vertical activity, and winter temperature experience of Greenland halibut Reinhardtius hippoglossoides in West Greenland waters. Marine Ecology Progress Series, vol. 508, 211-222.
- Box, J.E., and others (2009). Greenland ice sheet near-surface air temperature variability: 1840-2007, J. Climatology, vol. 22, 4029-4049, doi:10.1175/2009JCLI2816.1.
- Cantoni, C., and others (2020). Glacial drivers of marine biogeochemistry indicate a future shift to more corrosive conditions in an Arctic fjord. Journal of Geophysical Research: Biogeosciences, vol. 125, e2020JG005633. https://doi.org/10.1029/2020JG005633.
- Cape, Mattias, and others (2019). Nutrient release to oceans from buoyancy-driven upwelling at Greenland tidewater glaciers. Nature Geoscience, vol. 12, 34-39.
- Clarke, G., and others (2015). Projected deglaciation of western Canada in the twenty-first century. Nature Geoscience, vol. 8, 372-377. https://doi.org/10.1038/ngeo2407.
- Cooley, S.W., and others (2020). Coldest Canadian Arctic communities face greatest reductions in shorefast sea ice. Nature Climate Change, vol. 10, 533-538. https://doi.org/10.1038/s41558-020-0757-5.
- Cottier, F.R., and others (2010). Arctic fjords: a review of the oceanographic environment and dominant physical processes. Geological Society of London, vol. 344, 35-50.
- Cottier, F.R., and others (2005). Water mass modification in an Arctic fjord through cross-shelf exchange: the seasonal hydrography of Kongsfjorden, Svalbard. Journal of Geophysical Research: Oceans, 110:C12005.
- Crosswell, J.R., and others (2022). Geophysical controls on metabolic cycling in three Patagonian fjords. Progress in Oceanography, vol. 207. https://doi.org/10.1016/j.pocean.2022.102866.
- Cuevas, Antonio, and others (2019). Interplay between freshwater discharge and oceanic waters modulates phytoplankton size-structure in fjords and channel systems of the Chilean Patagonia. Progress in Oceanography, vol. 173, 103-113.
- Cui, X., and others (2022). Global fjords as transitory reservoirs of labile organic carbon modulated by organo-mineral interactions. Science Advances, vol. 8.
- Darelius, E. (2020). On the effect of climate trends in coastal density on deep water renewal frequency in sill fjords - A statistical approach. Estuarine, Coastal and Shelf Science, vol. 243. https://doi.org/10.1016/j.ecss.2020.106904.
- Del Bel Belluz, Justin, and others (2024). Phytoplankton community composition links to environmental drivers across a fjord to shelf gradient on the central coast of British Columbia. Frontiers in Marine Science. https://doi.org/10.3389/fmars.2024.1458677.
- Draganska-Deja, Katarzyna (2024). Characterization of suspended particles at different glacial bays at Spitsbergen. Oceanologia, vol. 6, 239-249. doi.org/10.1016/j.oceano.2023.12.001.
- Draganska-Deja, Katarzyna, and others (2024). Productivity of Spitsbergen fjords ecosystems in summer- Spatial changes of in situ primary production in Kongsfjorden and Hornsund in the period 1994 - 2019. Ecology and Evolution, vol. 14. https://doi.org/10.1002/ece3.11607.
- Ericson Y., and others (2019). Marine CO2 system variability in a high arctic tidewater-glacier fjord system, Tempelfjorden, Svalbard. Continental Shelf Research, vol. 181, 1-13. https://doi.org/10.1016/j.csr.2019.04.013.
- Farmer, D.M., and Freeland, H.J. (1983). The physical oceanography of Fjords. Progress in Oceanography, vol. 12, 147-220. https://doi.org/10.1016/0079-6611(83)90004-6.
- Faust, J.C., and others (2014). Terrigenous input to a fjord in central Norway records the environmental response to the North Atlantic Oscillation over the past 50 years. The Holocene, vol. 24, 1411-1418. https://doi.org/10.1177/0959683614544052.
- Faust, J.C., and others (2017). Geochemical characterization of northern Norwegian fjord surface sediments: A baseline for further paleo-environmental investigations. Continental Shelf Research, vol. 148, 104-115. https://doi.org/10.1016/j.csr.2017.08.015.
- Faust, J.C., and Knies, J. (2019). Organic matter sources in North Atlantic fjord sediments. Geochemistry, Geophysics, Geosystems, vol. 20, 2872-2885.
- Foss, Ø., and others (2024). Ocean warming drives immediate mass loss from calving glaciers in the high Arctic. Nature Communication, vol. 15. https://doi.org/10.1038/s41467-024-54825-7.
- Fransson, Agneta, and others (2015). Effect of glacial drainage water on the CO2 system and ocean acidification state in an Arctic tidewater-glacier fjord during two contrasting years. Journal of Geophysical Research: Oceans, 120. https://doi.org/10.1002/2014JC010320.
- Fransson, Agneta, and others (2016). Late winter-to-summer change in ocean acidification state in Kongsfjorden, with implications for calcifying organisms. Polar Biology, vol. 39, 1841-1857.
- Fransson, Agneta. and others (2020). Influence of glacial water and carbonate minerals on wintertime sea- ice biogeochemistry and the CO2 system in an Arctic fjord in Svalbard. Annals of Glaciology. https://doi.org/10.1017/aog.2020.52.
- Frey K.E., and others (2020). Arctic Ocean Primary Productivity: The Response of Marine Algae to Climate Warming and Sea Ice Decline. In Arctic Report Card 2020, R.L. Thoman, J. Richter-Menge, and M.L. Druckenmiller, eds. https://doi.org/10.25923/vtdn-2198.
- Galán, Alexander, and others (2021). Argo float reveals biogeochemical characteristics along the freshwater gradient off western Patagonia. Frontiers in Marine Science. doi:10.3389/fmars.2021.613265.
- Garreaud, René, and others (2009). Present-day south american climate. Paleogeography, Paleoclimatology, Paleoecology, vol. 281, 180-195. doi:10.1016/j.palaeo.2007.10.032.
- Garreaud, René, and others (2013). Large-scale control on the Patagonian climate. Journal of Climatology, vol. 26, 215-230. doi.org/ 10. 1175/JCLI-D- 12- 00001.1.
- Geertsma, M., and others (2022). The 28 November 2020 landslide, tsunami, and outburst flood -- A Hazard cascade associated with rapid deglaciation at Elliott Creek, British Columbia, Canada. Geophysical Research Letters, 49, e2021GL096716.
- Gerland, and others (2020). Long-term monitoring of landfast sea ice extent and thickness in Kongsfjorden, and related applications (FastIce). In Van den Heuvel, F., Hübner, C., Błaszczyk, M., Heimann, M., Lihavainen, H. eds., SESS Report 2019. Longyearbyen: Svalbard Integrated Arctic Earth Observing System: 160-167.
- Geyman, E.C., and others (2022). Historical glacier change on Svalbard predicts doubling of mass loss by 2100. Nature, 601, 374-379, https://doi.org/10.1038/s41586-021-04314-4, 2022.
- Giesbrecht, I.J.W., and others (2021). Watershed Classification Predicts Streamflow Regime and Organic Carbon Dynamics in the Northeast Pacific Coastal Temperate Rainforest - Giesbrecht - 2022 - Global Biogeochemical Cycles.
- González, Humberto, and others (2013). Land-ocean gradient in haline stratification and its effects on plankton dynamics and trophic carbon fluxes in Chilean Patagonian fjords. Progress in Oceanography, vol. 119, 32-47. http://dx.doi.org/10.1016/j.pocean.2013.06.003.
- González, H., and others (2019). Organic matter distribution, composition and its possible fate in the Chilean North-Patagonian estuarine system. Science of The Total Environment, vol. 657, 1419-1431. https://doi.org/10.1016/j.scitotenv.2018.11.445.
- Grønkjær, P, and others (2019). Feeding ecology of capelin (Mallotus villosus) in a fjord impacted by glacial meltwater (Godthåbsfjord, Greenland). Polar Biology, vol. 42, 81-98.
- Halbach, L., and others (2019). Tidewater glaciers and bedrock characteristics control the phytoplankton growth environment in an Arctic fjord. Frontiers in Marine Science, vol. 6, 254. https://doi.org/10.3389/fmars.2019.00254.
- Hanna, E., and others (2021). Greenland surface air temperature changes from 1981 to 2019 and implications for ice-sheet melt and mass-balance change. International Journal Climatology, vol. 41, 1336- 1352. https://doi.org/10.1002/joc.6771.
- Hare, A., and others (2020). Contrasting marine carbonate systems in two fjords in British Columbia, Canada: Seawater buffering capacity and the response to anthropogenic CO2 invasion. PLOS ONE. https://doi.org/10.1371/journal.pone.0238432.
- Hegseth, E.N., and others (2020). Phytoplankton seasonal dynamics in Kongsfjorden, Svalbard and the adjacent shelf. In Hop, H., Wiencke, C., eds., The ecosystem of Kongsfjorden, Svalbard. Cham, Switzerland: Springer: 173-227. Advances in Polar Ecology, vol. 2.
- Heide-Jørgensen, M.P., and others (2013). The significance of the north water polynya to arctic top predators. Ambio, vol. 42, 596-610. doi: 10.1007/s13280-012-0357-3.
- Heikkilä, M., and others (2022). Predicting the future of coastal marine ecosystems in the rapidly changing Arctic: the potential of palaeoenvironmental records. Anthropocene. https://doi.org/10.1016/j.ancene.2021.100319.
- Herbert, C., and others (2022). Tight benthic-pelagic coupling drives seasonal and interannual changes in iron-sulfur cycling in Arctic fjord sediments (Kongsfjorden, Svalbard). Journal of Marine Systems, vol. 225.
- Hodson, A., and others (2025). Methane in Svalbard. In Runge, and others, eds., SESS report 2024, Svalbard Integrated Arctic Earth Observing System, Longyearbyen, 106-137. https://doi.org/10.5281/zenodo.14425572.
- Hop, H., and others (2006). Physical and biological characteristics of the pelagic system across Fram Strait to Kongsfjorden. Progress in Oceanography, vol. 71, 182-231.
- Hop, H., and others (2020). Changes in sea-ice protist diversity with declining sea ice in the Arctic Ocean from the 1980s to 2010s. Frontiers in Marine Science, vol. 7, 243.
- Hoppe C.J.M., and others (2024). The effects of biomass depth distribution on phytoplankton spring bloom dynamics and composition in an Arctic fjord. Elementa: Science of the Anthropocene, vol. 12. https://doi.org/10.1525/elementa.2023.00137.
- Hopwood, Mark, and others (2018). Non-linear response of summertime marine productivity to increased meltwater discharge around Greenland. Nature Communications, vol. 9, 3256. doi:10.1038/s41467-018- 05488-8.
- Hopwood, Mark, and others (2020). Review article: How does glacier discharge affect marine biogeochemistry and primary production in the Arctic? The Cryosphere, vol. 14, 1-37.
- Hopwood, Mark, and others (2025). A close look at dissolved silica dynamics in Disko Bay, west Greenland. Global Biogeochemical Cycles, vol. 39, 2025.
- Hunt, B.P.V., and others (2024). Advancing an integrated understanding of land-ocean connections in shaping the marine ecosystems of Coastal Temperate Rainforest ecosystems. Limnology and Oceanography.
- Iriarte, José Luis, and others (2014). Oceanographic Processes in Chilean Fjords of Patagonia: from small to large-scale studies. Progress in Oceanography, vol. 129, 1-7.
- Iriarte, José Luis, and others (2007). Spatial and temporal variability of chlorophyll and primary productivity in surface waters of southern Chile (41.5 - 43S). Estuarine, Coastal and Shelf Science, 74, 471-480.
- Iriarte, José Luis (2018). Natural and Human Influences on Marine Processes in Patagonian Subantarctic Coastal Waters. Frontiers in Marine Science, vol. 5. https://doi.org/10.3389/fmars.2018.00360.
- Jackson, Jennifer, and others (2018). Warming From Recent Marine Heatwave Lingers in Deep British Columbia Fjord. Geophysical Research Letters, vol. 45, 9757-9764. https://doi.org/10.1029/2018GL078971.
- Jackson, Jennifer, and others (2022). Identification of a Seasonal Subsurface Oxygen Minimum in Rivers Inlet, British Columbia. Estuaries and Coasts, vol. 45, 754-771. https://doi.org/10.1007/s12237-021- 00999-y.
- Jackson, Jennifer, and others (2021). Deep Waters in British Columbia Mainland Fjords Show Rapid Warming and Deoxygenation from 1951 to 2020. Geophysical Research Letters, vol. 48, e2020GL091094. https://doi.org/10.1029/2020GL091094.
- Jacob, Barbara, and others (2014). Springtime size-fractionated primary production across hydrographic and PAR-light gradients in Chilean Patagonia (41-50°S). Progress in Oceanography, vol. 129, 75-84.
- Johannessen, S.C., and others (2019). Rain, Runoff, and Diatoms: the Effects of the North Pacific 2014-2015 Warm Anomaly on Particle Flux in a Canadian West Coast Fjord. Estuaries and Coasts, vol. 42, 1052-1065. https://doi.org/10.1007/s12237-018-00510-0.
- Johnsen, I.A, and others (2024). Coastal alterations influence deep water renewal in Norwegian sill fjords. Estuarine, Coastal and Shelf Science, vol. 297, 108604. https://doi.org/10.1016/j.ecss.2023.108604.
- Jones, E.M., and others (2020). Seasonal dynamics of carbonate chemistry, nutrients and CO2 uptake in a sub-Arctic fjord. Elemental Science of Anthropocene, vol. 8, 41.
- Juul-Pedersen, Thomas, and others (2015). Seasonal and interannual phytoplankton production in a sub- Arctic tidewater outlet glacier fjord, SW Greenland. Marine Ecology Progress Series, vol. 524, 27-38.
- Kohler, J., and others (2007). Acceleration in thinning rate on western Svalbard glaciers. Geophysical Research Letters, vol. 34. doi:10.1029/2007GL030681.
- Kristiansen, S., and others (2014). An Emiliania huxleyi dominated subsurface bloom in Samnangerfjorden, Western Norway. Importance of hydrography and nutrients. Sarsia, vol. 79, 357-368.
- Krupnik, I., and others (2010). SIKU: Knowing Our Ice. Springer, Dordrecht. https://doi.org/ 10.1007/978-90-481-8587-0.
- Laidre, K.L., and others (2008). Quantifying the sensitivity of Arctic marine mammals to climate-induced habitat change. Ecological Applications, vol. 18, 97-125.
- Leon-Muñoz, Jorge, and others (2018). Hydroclimatic conditions trigger record harmful algal bloom in western Patagonia (summer 2016). Scientific Reports, DOI: 10.1038/s41598-018-19461-4, 2018.
- León-Muñoz, Jorge, and others. Hydrographic shifts in coastal waters reflect climate-driven changes in hydrological regimes across Northwestern Patagonia. Scientific Reports, doi.org/10.1038/s41598-024- 71008-y, 2024.
- Linford, P., and others. Oceanographic processes driving low-oxygen conditions inside Patagonian fjords. Biogeosciences, vol. 21, 1433-1459. https://doi.org/10.5194/bg-21-1433-2024.
- Lydersen, C., and others (2014). The importance of tidewater glaciers for marine mammals and seabirds in Svalbard, Norway. Journal of Marine Systems, vol. 129, 452-471.
- Mankoff, K.D., and others (2021). Greenland ice sheet mass balance from 1840 through next week. Earth System Science Data, vol. 13, 5001-5025. DOI: 10.5194/essd-13-5001-2021.
- Mankoff, K.D., and others (2020). Greenland liquid water discharge from 1958 through 2019. Earth System Science Data, vol. 12, 2811-2841. DOI: 10.5194/essd-12-2811-2020.
- Marchese, C., and others (2022). Bioregionalization of the coastal and open oceans of British Columbia and Southeast Alaska based on Sentinel-3A satellite-derived phytoplankton seasonality. Frontiers in Marine Science, vol. 9. https://doi.org/10.3389/fmars.2022.968470.
- Matthews, J.B.L., and Heimdal, B.R. (1980). Pelagic productivity and food chain in fjord systems. In Fjord Oceanography, Freeland, H.J., Farmer, D.M., and Levings, C.D, eds. Springer, Boston, MA. https://doi.org/10.1007/978-1-4613-3105-6_34.
- Minowa, M., and others (2022). Effects of topography on dynamics and massloss of lake terminating glaciers in southern Patagonia. Journal of Glaciology. DOI: https://doi.org/10.1017/jog.2023.42.
- Navedo, Juan, and Vargas-Chacoff, Luis (2021). Salmon aquaculture threatens Patagonia. Science, vol. 372, 694.
- Nilsen, F., and others (2008). Fjord-shelf exchanges controlled by ice and brine production: the interannual variation of Atlantic Water in Isfjorden, Svalbard. Continental Shelf Research, vol. 28, 1838- 1853. https://doi.org/10.1016/j.csr.2008.04.015.
- Meire, M., and others (2017). Marine-terminating glaciers sustain high productivity in Greenland fjords. Global Change Biology, vol. 23, 5344-5357.
- Meire, Lorenz, and others (2023). Glacier retreat alters downstream fjord ecosystem structure and function in Greenland. Nature Geoscience, vol. 16, 671-674.
- Moon, T., and others (2017). Subsurface iceberg melt key to Greenland fjord freshwater budget. Nature Geoscience, vol. 11, 49-54. doi:10.1038/s41561- 017-0018-z.
- Morrison, J., and others (2011). A Method for Estimating Monthly Freshwater Discharge Affecting British Columbia Coastal Waters. Atmosphere-Ocean, vol. 50, 1-8. https://doi.org/10.1080/07055900.2011.637667.
- Oksman, M., and others (2022). Impact of freshwater runoff from the southwest Greenland Ice Sheet on fjord productivity since the late 19th century. The Cryosphere, vol. 16, 2471-2491.
- Oksman, M., and others (2024). Climate variability and glacier dynamics linked to fjord productivity changes over the last ca. 3300 years in Nuup Kangerlua, Southwest Greenland. Paleoceanography and Paleoclimatology.
- Pata, P.R., and others (2022). Persistent zooplankton bioregions reflect long-term consistency of community composition and oceanographic drivers in the NE Pacific. Progress in Oceanography, vol. 206. https://doi.org/10.1016/j.pocean.2022.102849.
- Pavlova, O., and others (2019). Changes in Sea-Ice Extent and Thickness in Kongsfjorden, Svalbard (2003-2016). In Hop, H. and Wiencke, C., eds. The Ecosystem of Kongsfjorden, Svalbard. Advances in Polar Ecology, Springer, Cham. https://doi.org/10.1007/978-3-319-46425-1_4.
- Pickard, G.L. (1961). Oceanographic Features of Inlets in the British Columbia Mainland Coast Journal of the Fisheries Board of Canada. https://doi.org/10.1139/f61-062.
- Pickard, G.L. (1973). Water structure in Chilean fjords. In Fraser, R., ed. Oceanography of the South Pacific. New Zealand National Commission for UNESCO Wellington.
- Pitcher, G.C., and others (2021). System controls of coastal and open ocean oxygen depletion. Progress in Oceanography, vol. 197. https://doi.org/10.1016/j.pocean.2021.102613.
- Quiñones, Renato, and others (2019). Environmental issues in Chilean salmon farming: a review. Reviews in Aquaculture, vol. 11, 375-402. https://doi.org/10.1111/raq.12337.
- Rebolledo, L., and others (2015). Late Holocene precipitation variability recorded in the sediments of Reloncaví Fjord (41°S, 72°W), Chile. Quaternary Research, vol. 84, 21-36. https://doi.org/10.1016/j.yqres.2015.05.006.
- Ribeiro, Sophia, and others (2017). Sea ice and primary production proxies in surface sediments from a High Arctic Greenland fjord: Spatial distribution and implications for palaeoenvironmental studies. AMBIO, vol. 46, 106.
- Ribeiro, Sophia, and others (2021). Vulnerability of the North Water ecosystem to climate change. Nature Communications, vol. 12, 4475.
- Rojas, N., and Silva, N. (2005). Early diagenesis and vertical distribution of organic carbon and total nitrogen in recent sediments from southern Chilean fjords (Boca del Guafo to Pulluche Channel). Investigaciones Marinas, vol. 33. https://doi.org/10.4067/S0717-71782005000200005.
- Rüggeberg, A., and others (2011). Water mass characteristics and sill dynamics in a subpolar cold-water coral reef setting at Stjernsund, northern Norway, Marine Geology, vol. 282, 5-12. https://doi.org/10.1016/j.margeo.2010.05.009.
- Saldías, Gonzalo, and others (2019). Freshwater structure and its seasonal variability off western Patagonia. Progress in Oceanography, vol. 174, 143-153. doi: 10.1016/j.pocean.2018.10.014.
- Schiøtt, S., and others (2022). Inuit and local knowledge on the marine ecosystem in Ilulissat Icefjord, Greenland. Human Ecology, 1-15.
- Sepúlveda, J., and others (2009). Late Holocene sea-surface temperature and precipitation variability in northern Patagonia, Chile (Jacaf Fjord, 44°S). Quaternary Research, vol. 72, 400-409. https://doi.org/10.1016/j.yqres.2009.06.010.
- Sepúlveda, J., and others (2011). Sources and distribution of organic matter in northern Patagonia fjords, Chile (~44-47°S): A multi-tracer approach for carbon cycling assessment. Continental Shelf Research, vol. 31, 315-329. https://doi.org/10.1016/j.csr.2010.05.013.
- Silva, Nelson, and others (2011). Land-ocean distribution of allochthonous organic matter in surface sediments of the Chiloé and Aysén interior seas (Chilean Northern Patagonia). Continental Shelf Research, vol. 31, 330-339. https://doi.org/10.1016/j.csr.2010.09.009.
- Skarðhamar J., and H. Svendsen (2005). Circulation and shelf-ocean interaction off North Norway. Continental Shelf Research, vol. 25, 1541-1560.
- Smeaton, C., and Austin, W.E.N. (2022). Understanding the role of terrestrial and marine carbon in the mid-latitude fjords of Scotland. Global Biogeochemical Cycles, vol. 36.
- Schmidt, C.E., and others (2025). The contrasting role of marine- and land-terminating glaciers on biogeochemical cycles in Kongsfjorden, Svalbard. Global Biogeochemical Cycles, vol. 39.
- Stenevik, E.K., and others (2008). Buoyancy and vertical distribution of Norwegian coastal cod (Gadus morhua) eggs from different areas along the coast. ICES Journal of Marine Science, vol. 65, 1198-1202.
- Svendsen, H., and others (2002). The physical environment of Kongsfjorden-Krossfjorden, an Arctic fjord system in Svalbard. Polar Research, vol. 21, 133-166.
- Syvitski, J.P.M., and others (1987). Fjords: Processes and Products. Springer-Verlag, N.Y.
- Sætre, R., and others (1988). Wind effects on the lateral extension of the Norwegian Coastal Water. Continental Shelf Research, vol. 8, 239-253.
- Skogseth, R., and others (2020). Variability and decadal trends in the Isfjorden (Svalbard) ocean climate and circulation - An indicator for climate change in the European Arctic. Progress in Oceanography, vol. 187. https://doi.org/10.1016/j.pocean.2020.102394.
- Smith, R.W., and others (2015). High rates of organic carbon burial in fjord sediments globally. Nature Geoscience, vol. 8, 450-453.
- Slater, D.A., and others (2015). Effect of near-terminus subglacial hydrology on tidewater glacier submarine melt rates. Geophysical Research Letters, vol. 42, 2861-2868.
- Straneo, F., and others (2022). An interdisciplinary perspective on Greenland's changing coastal margins. Oceanography, vol. 35, 106-117.
- Straneo, F., and others (2012). Characteristics of ocean waters reaching Greenland's glaciers. Annals of Glaciology, vol. 53, 202-210.
- Søreide, J.E., and others (2022). Seasonal mesozooplankton patterns and timing of life history events in high-arctic fjord environments. Frontiers in Marine Science. doi: 10.3389/fmars.2022.933461.
- Søreide, J.E., and others (2010). Timing of Blooms, Algal Food Quality and Calanus Glacialis Reproduction and Growth in a Changing Arctic. Global Change Biology, vol. 16, 3154-3163. doi:10.1111/j.1365-2486.2010.02175.x.
- Søreide, J.E., and others (2008). Seasonal feeding strategies of Calanus in the high-Arctic Svalbard region. Deep-Sea Research II, 55, 2225- 2244.
- Tommasi, D.A.G., and others (2013). The seasonal development of the zooplankton community in a British Columbia (Canada) fjord during two years with different spring bloom timing. Marine Biology Research, vol. 9, 129-144. https://doi.org/10.1080/17451000.2012.708044.
- Van Leeuwen, S.M., and others (2021). Climate change, marine resources and a small Chilean community: making the connections. Marine Ecology Progress Series, vol. 680: 223-246. https://doi.org/10.3354/meps13934.
- Vermassen, F., and others (2019). A reconstruction of warm-water inflow to Upernavik Isstrøm since 1925 CE and its relation to glacier retreat. Climate Past, vol. 15, 1171-1186. https://doi.org/10.5194/cp- 15-1171-2019.
- Vermassen, F., and others (2019). Evaluating ice-rafted debris as a proxy for glacier calving in Upernavik Isfjord, NW Greenland. J. Quaternary Science, vol. 34, 258-267. https://doi.org/10.1002/jqs.3095.
- Vernet, María, and others (2021). Spatial variability in rates of net primary production (NPP) and onset of the spring bloom in Greenland shelf waters. Progress in Oceanography, vol. 198.
- Vonnahme, T.R., and others (2023). Impact of winter freshwater from tidewater glaciers on fjords in Svalbard and Greenland; A review. Progress in Oceanography.
- Zimmermann F., and others (2023). Fine-scale spatial variation of northern shrimp and Atlantic cod across three Norwegian fjord systems and implications for management. Estuarine, Coastal and Shelf Science, vol. 292. https://doi.org/10.1016/j.ecss.2023.108435.
- Watts, E.G., and others (2024). Burial of organic carbon in Swedish Fjord sediments: Highlighting the importance of sediment accumulation rate in relation to fjord redox conditions. Journal of Geophysical Research-Biogeosciences, vol. 129.
- Wehrmann M., and others (2014). Iron and manganese speciation and cycling in glacially influenced high-latitude fjord sediments (West Spitsbergen, Svalbard): Evidence for a benthic recycling-transport mechanism Geochimica et Cosmochimica Acta, vol. 141, 628-655.
- Wolfe, A., and others (2015). Impact of advection loss due to wind and estuarine circulation on the timing of the spring phytoplankton bloom in a fjord. ICES Journal of Marine Science, vol. 73, 1589-1609. https://doi.org/10.1093/icesjms/fsv151.
- York A.V., and others (2020). Changes at the edge: trends in sea ice, ocean temperature and ocean color at the Northwest Atlantic/Southern Arctic interface. Annals of Glaciology, 1-15. doi:10.1017/aog.2020.66.