Hydrothermal vents and cold seeps
Writing team: Lisa A. Levin (coordinating author), Ana Colaço, Elva Escobar, Nadine Le Bris, Anna Metaxas, Paraskevi Nomikou, Samuel O. Popoola, Julia Sigwart, Paulo Sumida, Joshua Tuhumwire (lead member), Verena Tunnicliffe, Juying Wang (co-lead member), Hiromi Watanabe and Yadong Zhou.
Key points
- Discovery of new sites, novel habitats, new species and new chemosynthetic symbioses continues at a rapid pace for vents and seeps.
- Pressures are growing from climate change, fisheries and seabed mining, but protection for vents and seeps remains limited.
- New recognition, quantification and valuation of critical ecosystem services provided by vents and seeps heighten conservation imperatives.
- Inequities remain in knowledge and capacity transfer, particularly for Indigenous Peoples and low- and middle-income countries.
1. Introduction and context
Summary of baseline state provided by the first and second World Ocean Assessment
Hydrothermal vents and cold seeps host high microbial and animal biomass supported by chemosynthesis, involving microbe-animal symbioses, and contribute to broader productivity and fisheries in surrounding systems. They occur at mid-ocean ridges, volcanic arcs and hotspot-associated volcanic intraplate and back-arc spreading centres (vents) and on passive continental margins and subduction zones (seeps). Both vents and seeps exhibit high spatial and temporal variability of constraining environmental factors imposing physiological adaptations. Late successional systems including cold-water corals and inactive sulfide communities may persist over centuries. The high levels of endemism provide significant species discovery and provide model systems for understanding adaptation to extreme conditions and critical climate regulating services, as well as biotechnology and biomedical potential (e.g. Ref 25).
Technological advances have enabled discovery of thousands of sites in recent decades, as well as development of deep oil, gas and gas hydrate activities (seeps), and new mining exploration contracts (vents) issued in the Atlantic and Indian Oceans. Additional pressures include global warming, deoxygenation, ocean acidification and altered circulation, deep-sea fishing, waste dumping and plastic debris. There is growing recognition of the need for vent and seep protection. Conservation actions to date include scientific and industry codes of conduct, protected areas and International Union for the Conservation of Nature and Natural Resources Red List designations Ref 59.
Scope of the coverage
Recent regional exploration and discovery of vents and seeps are synthesized below, as well as key advances in knowledge and understanding of processes and threats, description of environmental change, social implications and knowledge and capacity gaps.
2. Environmental change since the second World Ocean Assessment
Changes in the overall status
Climate change
Rising ocean temperatures (see sect. 4, chap. 3) caused past gas hydrate destabilization Ref 38. The flux of methane from seabed seeps to the atmosphere is increasing in the Arctic and Antarctic Ref 114. In a warming ocean, present-day connectivity pathways may be disrupted due to shorter larval duration (thus smaller dispersal distance) and vertical position shifts Ref 83, changing current patterns (magnitude or direction) Ref 62 or loss of suitable habitat Ref 72.
Resource exploration and exploitation
Intense research occurs at some vent sites with likely impacts Ref 56. Recovery from a small, induced disturbance at a Mid-Atlantic Ridge vent found altered community structure after two years, with implications for deep-sea mining Ref 65. As of 2024, ISA has signed seven exploration contracts for vent deposits in the Atlantic and Pacific Oceans (see subsect. 5A, chap. 7). Exploration licenses are active in Japan, Tonga and Papua New Guinea, while Norway has initiated approvals. Exploitation of active vents may cause losses of species, functions and connectivity Ref 3 Ref 103 while knowledge of the ecological value of inactive/extinct sulphides is limited Ref 107. Offshore oil and gas development continues (see subsect. 5A, subchap. 3B), likely affecting associated methane seeps.
Bottom trawling (see subsect. 5A, subchap. 1C) exerts pressure on cold seep faunas as commercial species often congregate near seeps Ref 104. Foundation species were reduced at New Zealand seeps Ref 55 and trawl nets were tangled on carbonates on the Canadian Pacific Margin Ref 35. In Norway, broken carbonates and trawl marks had limited recolonization Ref 14. Hydrocarbon exploration on continental margins (see subsect. 5A, subchap. 3B) has located methane seep communities Ref 21. Extraction will likely diminish the methane supply that fuels chemosynthesis as observed by Boles and others (2023).
Factors contributing to the change observed
The changes are driven by rising populations and industry development increasing fishing pressure (see subsect. 5A, subchap. 1C), energy demand (see subsect. 5A, subchap. 3B) and emissions (causing warming) (see sect. 4, chap. 3), as well as rising metal demand and geostrategic contexts (see subsect. 5A, chap. 7), in part for electrification of transport.
Impacts and interactions of the change with other marine system components
Discoveries of large nursery areas of deep-sea species near vents and seeps (e.g. Ref 10) suggest that changes in these systems can influence recruitment to non-vent species, including from higher trophic levels.
Social, economic and cultural aspects associated with the change (since the second World Ocean Assessment)
As potential exploitation draws closer, societal awareness is growing, often with strong disagreements with State or ISA actions Ref 44. Mining at vents may intersect with sites of cultural heritage significance Ref 104 (see subsect. 5B, chap. 5).
Implications for the 2030 Agenda for Sustainable Development and for the Agreement under the United Nations Convention on the Law of the Sea on the Conservation and Sustainable Use of Marine Biological Diversity of Areas beyond National Jurisdiction
Protection supports Sustainable Development Goal 14 through sustainable management (target 14.2) and strengthening resilience using marine protection areas (MPAs) (Target 14.5). ISA (ISBA/27/C/8) declares support for Goal 14 through its regulatory framework for seabed mining, which requires trade-offs with Goal 7 (energy) and Goal 8 (industry). A draft regional environmental management plan (ISBA/27/C/38) reflects target 14.2 by classifying active hydrothermal vents as needing protection. Current mining code development aims to prevent environmental harm while mining in vent areas. Protection of vents and seeps, plus the ecosystem functions and services they provide, can help meet the Kunming-Montreal Global Biodiversity Framework targets. For the Agreement on Marine Biological Diversity of Areas beyond National Jurisdiction all four components are relevant.
3. Region-specific changes: new discoveries and new pressures
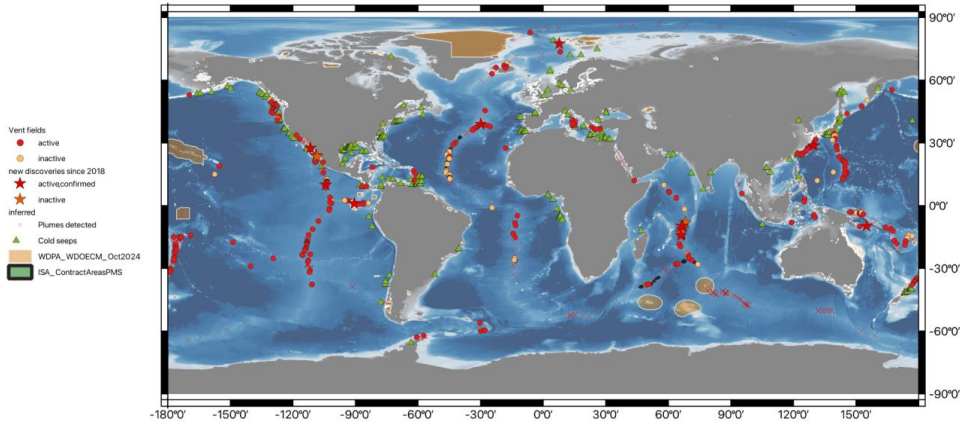
The figure below depicts previously known and recently discovered (since 2018) deep vents and seeps.
Figure Global map of confirmed active (red symbols) or reported inactive vent fields (orange symbols) and cold seeps (green triangles); for vents, stars denote vent fields discovered in 2018-2024. Brown polygons represent designated marine protected areas.
At shallow vents and seeps (less than 200 m; not mapped in the figure), species may coexist alongside photosynthetic algae and seagrass, and have greater exposure to anthropogenic changes including warming, ocean acidification, bottom fishing, pollution, or invasive species Ref 8. New technologies were increasingly used for analysing deep-sea biota, revealing magnificent vent assemblages Ref 85 Ref 99. Vent and seep faunas show heterogeneity linked to depth, surface production, oxygen minimum zones and geology Ref 94.
Arctic Ocean
First found in the 1990s, cold seep discovery continues in the Arctic Ref 95 Ref 96, as well as two recently discovered vent fields Ref 98 Ref 91 and many new vent species Ref 37 (see sect. 4, subchap. 5O).
North Atlantic Ocean, Baltic Sea, Black Sea, Mediterranean and North Sea
New vents were discovered on the Mid-Atlantic Ridge Ref 26. Other new vents discovered south of Iceland at the Reykjanes Ridge were noticeably lacking typical vent macrofauna Ref 100. In the northwest Atlantic margin, acoustic imaging revealed approximately 1,139 unique methane flare sites; most are shallower than 400 m, but some extend to 1400 m Ref 89. A newly discovered squat lobster in the Gulf of Mexico is the first west Atlantic seep endemic Ref 86.
South Atlantic Ocean and wider Caribbean
Seeps in the West Atlantic, the Caribbean and off Colombia are associated with linear diapir systems, salt diapirs resulting from salt tectonics, regions of mud volcanism and compact seafloor mounds; most knowledge is still associated with the oil and gas industry, with limited access to the science community and limited faunal studies Ref 2. Argentina's first methane seeps (approximately 500 m) are reported with dense infaunal assemblages Ref 21.
Indian Ocean, Arabian Sea, Bay of Bengal, Red Sea, Gulf of Aden and Persian Gulf
New discoveries include a rare serpentinite-hosted field Ref 61 and the first actively diffusing vent system in the Red Sea Ref 106; both host unique microbial communities without typical vent macrofauna. Faunal studies described over 30 new taxa and indicate within-region biogeographic discontinuity (e.g. Ref 52). Two new seep communities were reported in the northern Indian Ocean with biogeographic affiliations with the West Pacific vent/seep communities Ref 66, and a new active cold seep site occurs in the Bay of Bengal Ref 92.
North Pacific Ocean
Serpentine-hosted seep communities at mud volcanoes on the Mariana Forearc, where methane-rich fluids are highly alkaline (pH over 12), support many novel species Ref 28 (see sect. 4, subchap. 5L). New northeast Pacific Ocean vent and seep sites continue to be discovered Ref 41 notably including 1,600 methane flares mapped in the Canadian Pacific Ref 35 and eight new seeps off the coast in Oregon and Washington Ref 94. Low temperature ridge-flank hydrothermal springs in the northeast Pacific Ocean support deep-sea octopus nurseries Ref 10.
South Pacific Ocean and equatorial Pacific Ocean
A newly discovered vent site in the Southwest Pacific may contribute to faunal connectivity Ref 19. Other new vent sites in the Galapagos Rift expand the records of vent species Ref 30.
Southern Ocean
Three new deep-sea vent sites have been reported in the Atlantic region of the Southern Ocean Ref 63. Major new seepage sites in the Ross Sea indicate likely widespread climate-sensitive phenomena around Antarctica Ref 93.
4. Remaining key knowledge and capacity gaps and new gaps
Advances in technology
Many technological advances have improved understanding of chemosynthetic ecosystems, such as: (i) evolving molecular tools to support systematics and studies of adaptations to extreme environments and genetic connectivity (e.g. Ref 115); (ii) photogrammetry methods for 3D reconstructions to aid habitat change studies (e.g. Ref 105); (iii) neural network approaches to image classification Ref 109; and, (iv) novel computer algorithms for seep identification Ref 89.
Novel habitats and processes
Important new findings include subseafloor vent faunal habitats Ref 23 and off-axis hydrothermalism Ref 67 and novel chemo-symbioses Ref 47 and inactive vent species Ref 29. Microbial processes are implicated in primary productivity at inactive vent deposits Ref 1, in carbon cycling and nutrient export Ref 110 and in trophic networks in vent communities Ref 73. A comprehensive genomic catalogue of microbes from seeps Ref 50 enables exploration of functional processes such as biogeochemical cycling.
Dynamics, succession and sphere of influence
Long-term observations and experiments reveal high stability and slow recovery of vent ecosystems in slow-spreading ridges following disturbance Ref 65. Evidence that vents and seeps affect the ocean outside their perimeters accumulates, such as iron from rising vent plumes inducing phytoplankton blooms in the Southern Ocean Ref 5 and enhanced faunal abundance on the seafloor Ref 57. At seeps, input from both chemosynthesis and photosynthesis support distinctive infaunal communities in an intermediate zone between methane seeps and background Ref 7.
Stepping stones
Population connectivity could enhance resilience to disturbance. Highly divergent gene flow directions and ranges exist among closely allied vent species Ref 69. A novel, but isolated, Arctic vent ecosystem in a region with imminent mineral exploration provides evidence for historical connectivity Ref 37. Range analyses divide the Indian Ocean into three vent provinces that suggest separate management areas Ref 116. In the northwest Pacific Ocean, the Okinawa Trough has high endemicity Ref 24 while connectivity along the volcanic arcs appears limited for many species Ref 43. Most vent species are known only within one limited region; for a few species, the Manus Basin is the key region for cross-equator linkages Ref 24. At both vents and seeps, some widespread species form localized genetic units with rare long-distance dispersal events Ref 54.
Ecosystem services
Hydrothermal vents provide regulating services such as biogeochemical cycling of manganese, iron and nitrogen and carbon fixation. Provisioning services include marine genetic resources Ref 45 (see subsect. 5A, chap. 5) and potential metals from mining (see subsect. 5A, chap. 7). Supporting services include nutrient transfers to deep-sea and nursery areas at both active and inactive vents. Cultural values flow from scientific research (e.g. models for extraterrestrial life) and creative inspiration Ref 33. Cold seeps provide provisioning services through commercial fish/crustacean species and regulating services through carbon sequestration Ref 58. This new understanding heightens conservation imperatives.
Consequences of environmental impacts, governance challenges, conservation status
About half of known vent sites fall in areas beyond national jurisdiction where many are under mining exploration contracts; regulations on the conservation status of vents within contract areas are under development. Of all other vents, 17% are covered by non-targeted conservation interventions while 8% benefit from vent-specific actions Ref 68. For new and updated areas, see the table below. Four southwest Pacific States initially declared a moratorium on deep-sea mining (Ref 70); today 32 States have taken a position against deep-seabed mining in international waters. Thomas and others (2021) find the limited distribution of most vent molluscs is the key vulnerability, with notable extinction threat in mining contract areas. However, regular distribution assessment updates are needed. Conservation is in place for very few cold seeps although many are vulnerable to anthropogenic disturbance and climate change Ref 58 Ref 14. An economic valuation identified highest willingness to pay to conserve Costa Rican seeps with endemic species Ref 80. Governance in the areas beyond national jurisdiction is problematic due to fragmented and variable approaches Ref 68, and protections remain limited.
Table Emerging threats, risks and conservation measures since 2019
| Basin | Threat actions | Other risks | Conservation measures | Citations |
|---|---|---|---|---|
| Arctic | Norway approved (but paused in Dec. 2024) exploration for extraction of polymetallic sulphides | Warming waters could bring colonists to seeps and vents northward challenging endemic species | Delivorias, A. 2024. Åström, E.K. and others, 2020 | |
| North Atlantic and Mediterranean | Exploration of three ISA contract areas in progress in active vent sites | Bottom contact fishing on shallower vent areas. Marine litter (mostly plastics) in vent habitats from abandoned fishing gear | Luso hydrothermal field MPA in Portugal. ISA considers regional environmental management plan proposal to protect all active vents from mining in contract areas | Governo dos Açores. 2022 ISA, 2022 Consoli, P. and others, 2021 |
| Western tropical Atlantic and Caribbean | Submarine slides caused by methane hydrate destabilization in a warming ocean. Lack of literacy and capacity to address impacts | Ruffine, L. and others, 2023 Amon D. and others, 2022 | ||
| South Atlantic | Oil and gas exploration in deep water continues off Argentina near newly discovered seeps | Ascension Island MPA; (United Kingdom). Covers 750 km of ridge crest with known vent sites | Ascension Island Government, 2021 Bravo M. and others, 2024 | |
| Northeast Pacific | Notable climate-induced deoxygenation in the MPA Ref 87 may affect shallow vents and larvae | Bottom contact fishing on shallower vent areas. Deep seabed mining could affect vents | Tang. Gwan-hačxwiqak-Tsigis MPA now including all confirmed hydrothermal vents in Canada and some methane seeps | Canada Gazette, 2024 Ross, T. and others, 2020 |
| Northwest Pacific | Microplastic accumulation in cold seep animals | Northern part of Central (Naka) Mariana Ridge and West Mariana Ridge MPA, Japan. Includes three vent sites | Teng, J. and others, 2023 Watanabe H.K. and Yamakita T, 2022 Government of Japan, 2020. | |
| Southwest Pacific | Potential renewal of seabed exploitation licence for polymetallic sulphides at Solwara, Manus Basin, Papua New Guinea | Monga Bay, 2024 | ||
| Indian Ocean | Exploration of four ISA mining contract areas in progress at active vent sites | Microplastic accumulation in vent animals | Park, B. and others, 2024 | |
| Global | Seabed mining | Ocean warming and acidification | Assessment of hydrothermal vent species against the International Union for Conservation of Nature (IUCN) and Natural Resources Red List criteria. Of these, 114 fall into the vulnerable, endangered or critically endangered categories | Thomas, E.A. and others, 2021 |
Source: Prepared by the writing team
Key remaining knowledge gaps
Many knowledge gaps identified in the second World Ocean Assessment Ref 60 remain, including the distribution, biogeography and connectivity of vent and seep biota, resilience and time and space scales of recovery from disturbance (trawling, mining, climate change) and vent and seep roles in biogeochemical cycles. Capacity gaps are being identified Ref 13 and addressed Ref 53, but limited access to funding, research vessels, low-cost instruments and taxonomic expertise remain, creating global inequity. Small island developing States, for example, although often in close proximity, have limited access to hydrothermal vents and seeps. Currently several international programmes offer training opportunities and philanthropic foundations offer some research vessel support.
Newly recognized knowledge gaps
Regional gaps in location of new sites and in novel habitats, functions, symbioses and other mutualistic interactions among species, and genetic resource potential at vents and seeps exist. Additional key gaps include vent-seep-organic fall connectivity and stepping-stone functions including the role of inactive and ecotone/transition vent Ref 15 Ref 74 and seep Ref 79 sites, and animal-microbe interactions at active vents Ref 73, seeps Ref 80 and inactive sites.
References
- Achberger, Amanda M., and others (2024). Inactive hydrothermal vent microbial communities are important contributors to deep ocean primary productivity. Nature Microbiology, vol. 9, No. 3, pp. 657- 668.
- Aguilar Pérez, Maria Isabel, and others (2024). Review of cold seeps in the Western Atlantic, focusing on Colombia and the Caribbean. Frontiers in Marine Science, vol. 11, art. 1430377.
- Alfaro-Lucas, Joan, and others (2024). High functional variability across the world's deep-sea hydrothermal communities. Proceedings of the National Academy of Sciences of the United States of America., vol. 121, art. e2403899121.
- Amon, Diva, and others (2022). My Deep Sea, My Backyard: a pilot study to build capacity for global deep-ocean exploration and research. Philosophical Transactions of the Royal Society B, vol. 377, art. 20210121.
- Ardyna, Mathieu, and others (2019). Hydrothermal vents trigger massive phytoplankton blooms in the 691 Southern Ocean. Nature Communications, vol. 10, art. 2451.
- Ascension Island Government (2021). The Ascension Island Marine Protected Area Management Plan 2021-26. Ascension Island Government Conservation and Fisheries Directorate.
- Ashford, Oliver, and others (2021). A chemosynthetic ecotone - 'chemotone' - in the sediments surrounding deep-sea methane seeps. Limnology and Oceanography. vol. 66, No. 5, pp. 1687-1702.
- Åström, Emmelie K. L., and others (2018). Methane cold seeps as biological oases in the high-Arctic deep sea. Limnology and Oceanography, vol. 63, pp. S209-S231.
- Åström, Emmelie K. l. and others (2020). Cold seeps in a warming Arctic: Insights for benthic ecology. Frontiers in Marine Science, vol 7, art. 244.
- Barry, James P., and others (2023). Abyssal hydrothermal springs-Cryptic incubators for brooding octopus. Science Advances, vol. 9, No. 34, art. eadg3247.
- Baumberger, Tamara, and others (2018). Mantle-derived helium and multiple methane sources in gas bubbles of cold seeps along the Cascadia Continental Margin. Geochemistry, Geophysics, Geosystems, vol. 19, pp. 4476-4486.
- Beaulieu, Stace E., and Kamil M. Szafranski (2020). InterRidge Global Database of Active Submarine Hydrothermal Vent Fields. Version 3.4 PANGAEA. https://doi.pangaea.de/10.1594/PANGAEA.917894.
- Bell, Katherine L.C., and others (2023). Exposing inequities in deep-sea exploration and research: results of the 2022 Global Deep-Sea Capacity Assessment. Frontiers in Marine Science. vol. 10, art. 1217227.
- Bellec, Valerie Karin, and others (2024). New cold seep sites on the continental slope southwest to Svalbard. Frontiers in Earth Science, vol. 12, art.1328357.
- Boschen, Rachel, and Ana Colaco (2021). Northern Mid-Atlantic Ridge hydrothermal habitats: A systematic review of knowledge status for environmental management. Frontiers in Marine Science, vol. 8, art. 657358.
- Bohrman, Gerhard (2019). The Expedition PS119 of the Research Vessel Polarstern to the Eastern Scotia Sea in 2019. Berichte zur Polar-und Meeresforschung, Reports on polar and marine research, vol. 736.
- Bohrmann, Gerhard, and others (2024). Discovery of the first hydrothermal field along the 500-km-long Knipovich Ridge offshore Svalbard (the Jøtul field). Scientific Reports, vol. 14, art. 10168.
- Boles, James R., and others (2023). Hydrocarbon production reduces natural methane seeps in the Santa Barbara channel. Marine and Petroleum Geology, vol, 151, art. 106187.
- Boulart, Cedric, and others (2022). Active hydrothermal vents in the Woodlark Basin may act as dispersing centres for hydrothermal fauna. Communications Earth & Environment, vol. 3, No. 1, p. 64.
- Bravakos, Panos, and others. Genomic adaptation of Pseudomonas strains to acidity and antibiotics in hydrothermal vents at Kolumbo submarine volcano, Greece. Sci Rep vol. 11, 1336, 2021.
- Bravo, Maria Emilia, and others (2024). Discovery of deep-sea cold seeps from Argentina host singular trophic linkages and biodiversity. Deep-sea Research Part I, vol. 211, art. 104361.
- Breusing, Corinna., J., and others (2023). Differential patterns of connectivity in Western Pacific hydrothermal vent metapopulations: A comparison of biophysical and genetic models. Evolutionary Applications, vol. 16, No. 1, pp. 22-35.
- Bright, Monica, and others (2024). Animal life in the shallow subseafloor crust at deep-sea hydrothermal vents. Nature Communications, vol. 15, art. 8466.
- Brunner, Otis, and others (2022). Species assemblage networks identify regional connectivity pathways among hydrothermal vents in the Northwest Pacific. Ecology and Evolution, vol. 12, No. 12, art. e9612.
- Bruno, Renato, and others (2019). Worms' antimicrobial peptides. Marine Drugs, vol. 17, No. 512.
- Butterfield, David A., and others (2023). Discovery of three hydrothermal vent fields on the Mid-Atlantic Ridge with Falkor (too). AGU Fall Meeting Abstracts vol. 2023, No. 1281, p. OS11B-1281.
- Canada Gazette (2024). Part II, Volume 158, Number 13. https://gazette.gc.ca/rp-pr/p2/2024/2024-06-19/html/sor-dors122-eng.html.
- Chen, Chong, and others (2023). Unexpected discovery of a serpentinite-hosted chemosynthetic ecosystem on Asùt Tesoru Seamount, Mariana Forearc. Marine Ecology, vol. 44, No. 5, art. e12759.
- Chen, Chong, and others (2024a). Serpentinite-hosted chemosynthetic community of South Chamorro Seamount, Mariana Forearc. Marine Ecology, vol. 45 art. e12808.
- Chen, Chong, and others (2024b). Hydrothermal vent fauna of the Galápagos Rift: updated species list with new records. Marine Biodiversity, vol. 54, No. 2, art.16.
- ChEssBase, Ocean Biogeographic Information System: ChEssBase (CoML) (2019). Available at http://www.gbif.org/dataset/83b90d28-f762-11e1-a439-00145eb45e9a. Accessed on 2015-05-06.
- Consoli, Pierpaolo, and others (2021). Marine litter pollution associated with hydrothermal sites in the Aeolian archipelago (western Mediterranean Sea). Science of the Total Environment, vol. 773, art. 144968.
- Deep Ocean Stewardship Initiative (DOSI) (2023). Hydrothermal Vent Ecosystems - Information Sheet. Deep-Ocean Stewardship Initiative. https://www.dosi-project.org/wp-content/uploads/vents-ecosystems-info-sheet.pdf.
- Delivorias, A. (2024). Norway to mine part of the Arctic seabed. PE 757.616, EPRS. https://www.europarl.europa.eu/RegData/etudes/ATAG/2024/757616/EPRS_ATA(2024)757616_EN.pdf.
- Department of Fisheries and Oceans (DFO) (2018). Assessment of Canadian Pacific Cold Seeps against Criteria for Determining Ecologically and Biologically Significant Areas. DFO Can. Sci. Advis. Sec. Sci. Resp. 2018/002.
- Dueñas Luisa F., and others (2021). Fauna associated with cold seeps in the deep Colombian Caribbean. Deep-Sea Research vol. 173, art. 103552.
- Eilertsen, Mari H., and others (2024). Diversity, habitat endemicity and trophic ecology of the fauna of Loki's Castle vent field on the Arctic Mid-Ocean Ridge. Scientific Reports, vol. 14, No. 1, art. 103.
- El bani Altuna, N., and others (2021). Deglacial bottom water warming intensified Arctic methane seepage in the NW Barents Sea. Communications, Earth and Environment, vol. 2, art. 188.
- Etiope, Giuseppe, and others (2019). Gridded maps of geological methane emissions and their isotopic signature. Earth System Science Data, vol. 11, No. 1, pp. 1-22.
- Feng, Dong, and others. Cold seep systems in the South China Sea: An overview. Journal of Asian Earth Sciences, vol. 168, pp. 3-16, 2018.
- Galkin, Sergei V., and others (2019). Comprehensive research of ecosystems of hydrothermal vents and cold seeps in the Bering Sea (Cruise 82 of the R/V Akademik MA Lavrentyev). Oceanology, vol. 59, pp. 618-62.
- Gerdes, Klaas, and others (2021). Megafauna of the German exploration licence area for seafloor massive sulphides along the Central and South East Indian Ridge (Indian Ocean). Biodiversity Data Journal, vol. 9, art. e69955.
- Giguère Thomas, N., and Verena Tunnicliffe (2021). Beta diversity differs among hydrothermal vent systems: Implications for conservation. PLoS ONE, vol. 16, art. e0256637.
- Gilbert, Natasha (2024). First approval for controversial sea-bed mining worries scientists. Nature, vol. 625, pp. 435-436.
- Giordano, Daniela (2020). Bioactive molecules from extreme environments. Marine drugs, vol. 18, No. 12, art.640.
- Goffredi, Shana K., and others (2021). Mixotrophic chemosynthesis in a deep-sea anemone from hydrothermal vents in the Pescadero Basin, Gulf of California. BMC Biology, vol. 19, art. 8.
- Goffredi, Shana K., and others (2020). Methanotrophic bacterial symbionts fuel dense populations of deep-sea feather duster worms (Sabellida, Annelida) and extend the spatial influence of methane seepage. Science Advances, vol. 6, art. eaay8562.
- Government of Japan. Ministry of the Environment (2020). Designation of offshore seabed natural environment conservation areas. https://www.env.go.jp/press/108741.
- Governo dos Açores (2019). Portaria n.o 68/2019, de 26 de setembro - aprova o Regulamento para o exercício da pesca na zona marítima do campo hidrotermal LUSO.
- Han, Yingchun, and others (2023). A comprehensive genomic catalog from global cold seeps. Scientific Data, vol. 10, art. 596.
- International Seabed Authority (2022). Regional environmental management plan for the Area of the northern Mid-Atlantic Ridge with a focus on polymetallic sulphide deposits. ISBA/27/C/38, 56 pp.
- Jang, Sook-Jin, and others (2023). Geographical subdivision of Alviniconcha snail populations in the Indian Ocean hydrothermal vent regions. Frontiers in Marine Science, vol. 10, pp. 1139190.
- Johannes Kristen N., and others (2023). Seafloor observation scenario exploration tool enabling representative exploration of the global deep seafloor. Frontiers in Marine Science, vol. 10, art. 251562.
- Jollivet Didier, and others (2024). Preferential pathways of dispersal and role of the AMOC in connectivity. Ref. WP1. iAtlantic Deliverable 1.5. iAtlantic.
- Jones, Mckenzie Margaret (2023). Global and Regional Variations in Methane Seep Communities MSc Thesis, Victoria University of Wellington. 118 pp.
- Juniper, S. Kim, and others (2019). Remote monitoring of a deep-sea marine protected area: The Endeavour Hydrothermal Vents. Aquatic Conservation: Marine and Freshwater Ecosystems, vol. 29, pp. 84-102.
- Klunder, Lise, and others (2020). A molecular approach to explore the background benthic fauna around a hydrothermal vent and their larvae: Implications for future mining of deep-Sea SMS deposits. Frontiers in Marine Science, vol. 7, 10.3389/fmars.2020.00134.
- Le, Jennifer Tran., and others (2022). Using deep-sea images to examine ecosystem services associated with methane seeps, Marine Environmental Research, vol. 181, art. 105740.
- Le Bris, Nadine, and others (2017). The First World Ocean Assessment - WOA I_Chapter 45. Hydrothermal vents and cold seeps.
- Le Bris, Nadine and others (2021). The Second World Ocean Assessment - WOA II_Chapter 7P: Hydrothermal vents and cold seeps.
- Lecoeuvre, Aurelien, and others (2021). Microbial ecology of the newly discovered serpentinite-hosted Old City hydrothermal field (southwest Indian ridge). The ISME journal, vol. 15, No. 3, art. 818-832.
- Levin Lisa A, and others (2020). Climate change considerations are fundamental to management of deep-sea resource extraction. Global Change Biology, vol. 26, art. 4664.
- Linse, Katrine, and others (2022). Megabenthos habitats influenced by nearby hydrothermal activity on the Sandwich Plate, Southern Ocean. Deep Sea Research Part II: Topical Studies in Oceanography, vol. 198, art. 105075.
- MacDonald, Ian R., and others (2020). The asphalt ecosystem of the Southern Gulf of Mexico: Abyssal habitats across space and time. In Murawski, S., and others, Scenarios and Responses to Future Deep Oil Spills. Springer, Cham, pp. 132-146.
- Marticorena, Julien, and others (2021). Recovery of hydrothermal vent communities in response to an induced disturbance at the Lucky Strike vent field (Mid-Atlantic Ridge). Marine Environmental Research, 168, art. 105316.
- Mazumdar, Asis, and others (2019). The first record of active methane (cold) seep ecosystem associated with shallow methane hydrate from the Indian EEZ. Journal of Earth System Science, vol. 128, pp.1-8.
- McDermott, Jill M., and others (2022). Discovery of active off-axis hydrothermal vents at 9 54' N East Pacific Rise. Proceedings of the National Academy of Sciences, vol. 119, No. 30, art. e2205602119.
- Menini, Elisabetta, and others (2023). Towards a global strategy for the conservation of deep-sea active hydrothermal vents. npj Ocean Sustainability, vol. 2, No. 1, p. 22.
- Methou, Pierre, and others (2024). Genetic connectivity and isotopic niches of alvinocaridid shrimps from chemosynthetic habitats in Aotearoa/New Zealand, with a new Alvinocaris species. Marine Ecology Progress Series, vol. 739, pp. 85-109.
- MSG Secretariat (2020). https://msgsec.info/wp-content/uploads/documentsofcooperation/2023-Aug-24-UDAUNE-DECLARATION-on-Climate-Change-by-Members-of-MSG.pdf.
- Monga Bay (2024). https://news.mongabay.com/2024/09/not-merely-exploration-png-deep-sea-mining-riles-critics-surprises-officials/.
- Morato, Telmo, and others (2020). Climate-induced changes in the suitable habitat of cold-water corals and commercially important deep-sea fishes in the North Atlantic. Global Change Biology, vol. 26, art. 2181.
- Murdock, Sheryl A., and others (2021). Emergent "core communities" of microbes, meiofauna and macrofauna at hydrothermal vents. ISME Communication vol. 1, art 27.
- Neufeld, Monika and others (2022). Non-vent megafaunal communities on the Endeavour and Middle Valley segments of the Juan de Fuca Ridge, Northeast Pacific Ocean. Frontiers in Marine Science, vol. 9, art. 849976.
- Olu, Karine, and others (2010). Biogeography and potential exchanges among the Atlantic Equatorial Belt cold-seep faunas. PLOS One, vol. 5, art. e11967.
- Paduan, Jennifer B., and others (2018). Discovery of hydrothermal vent fields on Alarcón Rise and in southern Pescadero Basin, Gulf of California. Geochemistry, Geophysics, Geosystems, vol. 19, pp. 4788-4819.
- Park, Byeongyong, and others (2024). Microplastic contamination of a benthic ecosystem in a hydrothermal vent. Environmental Science and Technology, vol. 58, pp. 7636-7642.
- Parsons Patrice, K.F., and others (2024). Hydrothermal seepage of altered crustal formation water seaward of the Middle America Trench, offshore Costa Rica. Geochemistry, Geophysics, Geosystems, vol. 25, art. e2023GC011246.
- Pereira, Olívia Soares, and others (2021). The dynamic influence of methane seepage on macrofauna inhabiting authigenic carbonates. Ecosphere vol. 12, No. 10, art 303744.
- Pereira, Olivia Soares, and others (2024). Understanding and valuing human connections to deep-sea methane seeps off Costa Rica. Ecological Economics, vol 223, art. 108228.
- Pereira, Olivia Soares (2024). Diversity and trophic dynamics, resilience, and value of hardground invertebrates at methane seeps and on mineral-rich slopes in the deep Eastern Pacific. PhD Dissertation. University of California, San Diego., La Jolla, California.
- Quattrini, Andrea M., and others (2015). Exploration of the canyon-incised continental margin of the Northeastern United States reveals dynamic habitats and diverse communities. PLoS ONE vol. 10, No. 10, art. e0139904.
- Rakka, Maria, and others (2021). Embryo and larval biology of the deep-sea octocoral Dentomuricea aff. meteor under different temperature regimes. Peer J, vol. 9.
- Rarez-Llodra, Eva, and others (2010). Deep, diverse and definitely different: unique attributes of the world's largest ecosystem, Biogeosciences, vol. 7, art. 2851-2899.
- Ramirez-Llodra, Eva, and others (2024). The emerging picture of a diverse deep Arctic Ocean seafloor: From habitats to ecosystems. Elementa: Science of the Anthropocene, vol. 12, No. 1.
- Rodriguez-Flores, Paula Carolina, and others (2024). Integrative taxonomy reveals a new species of deep-sea squat lobster (Galatheoidea, Munidopsidae) from cold seeps in the Gulf of Mexico. Zoosystems Evolution, vol. 100, pp. 1243-1257.
- Ross, Tetyana, and others (2020). Rapid deep ocean deoxygenation and acidification threaten life on Northeast Pacific seamounts. Global Change Biology, vol. 26, No. 11, pp. 6424-6444.
- Ruffine, Livio, and others (2023). Environmental challenges related to methane hydrate decomposition from climate change scenario and anthropic activities: State of the art, potential consequences and monitoring solutions. Earth-Science Reviews, vol. 246, art. 104578.
- Ruppel, Carolyn D., and others (2024). Methane seeps on the U.S. Atlantic margin: An updated inventory and interpretative framework. Marine Geology, vol. 471, art. 107287.
- Ryu, Taewoo, and others (2019). The first reference transcriptome assembly of the stalked barnacle, Neolepas marisindica, from the Onnuri Vent Field on the Central Indian Ridge. Marine Genomics, vol. 48, art. 100679.
- Sahlström, Fredrik, and others (2023). Mineralogical distribution and genetic aspects of cobalt at the active Fåvne and Loki's Castle seafloor massive sulfide deposits, Arctic Mid-Ocean Ridges. Ore Geology Reviews, vol. 153, art. 105261.
- Sangodkar, Nitisha, and others (2023). Macrofaunal distribution, diversity, and its ecological interaction at the cold seep site of Krishna-Godavari Basin, East Coast of India. Microbial Ecology, vol. 85, No. 1, arts. 61-75.
- Seabrook, Sarah, and others (2023). Emergent Antarctic seafloor seeps: A tipping point reached? ResearchSquare. DOI: https://doi.org/10.21203/rs.3.rs-3657723/v1.
- Seabrook, Sarah, and others (2024). Ubiquitous but unique: Water depth and oceanographic attributes shape methane seep communities. Limnology and Oceanography, vol. 69, No. 5, pp. 1218-1232.
- Sen, Arunima, and others (2019). Atypical biological features of a new cold seep site on the Lofoten-Vesterålen continental margin (northern Norway). Scientific Reports, vol. 9, No. 1, art. 1762.
- Serov, Pavel, and others (2024). Geological and glaciological controls of 21,700 active methane seeps in the northern Norwegian Barents Sea. Frontiers in Earth Science, vol. 12, art. 1404027.
- Soule, Samuel Adam, and others (2018). Exploration of the northern Guaymas Basin. Oceanography, vol. 31, No. 1, pp. 39-41.
- Stensland, Anne, and others (2019). He along the ultraslow spreading AMOR in the Norwegian-Greenland Seas. Deep Sea Research Part I: Oceanographic Research Papers, vol. 147, pp. 1-11.
- Sun, Jin, and others (2020). Nearest vent, dearest friend: biodiversity of Tiancheng vent field reveals cross-ridge similarities in the Indian Ocean. Royal Society Open Science, vol. 7, No. 3, art. 200110.
- Taylor James, and others (2021). The discovery and preliminary ecological and faunal descriptions of three new Steinaholl vent sites, Reykjanes Ridge, Iceland. Frontiers in Marine Science, vol. 8 art. 520713.
- Teng, Jia, and others (2023). First record of microplastics in Gigantidas platifrons (Mytilidae: Bathymodiolus) and Shinkaia crosnieri (Munidopsidae: Shinkaia) from a cold-seep in the South China Sea. Marine Pollution Bulletin, vol. 185, art. 114523.
- Thomas, Elin A., and others (2021). A global Red List for hydrothermal vent molluscs. Frontiers in Marine Science, vol. 8, art. 13022.
- Tunnicliffe, Verena and others (2023). Hydrothermal vent fauna of the western Pacific Ocean: Distribution patterns and biogeographic networks. Diversity and Distributions, vol. 30, No. 2, art. 313794.
- Turner, Phillip, J., and others (2020). Methane seeps on the US Atlantic margin and their potential importance to populations of the commercially valuable deep-sea red crab, Chaceon quinquedens. Frontiers in Marine Science, vol.7, art.75.
- Van Audenhaege, Loïc., and others (2024). Monitoring ecological dynamics on complex hydrothermal structures: A novel photogrammetry approach reveals fine-scale variability of vent assemblages. Limnology and Oceanography, vol. 69, No. 2, pp. 325-338.
- Van der Zwan, Froukje M., and others (2023). Widespread diffuse venting and large microbial iron-mounds in the Red Sea. Communications Earth & Environment, vol. 4, No. 1, art.496.
- Van Dover, Cindy L., and others (2020). Research is needed to inform environmental management of hydrothermally inactive and extinct polymetallic sulfide (PMS) deposits. Marine Policy, 121, art. 104183.
- Van Putten, E. Ingrid, and others (2023). History matters: societal acceptance of deep-sea mining and incipient conflicts in Papua New Guinea. Maritime Studies, vol. 22, No. 3, art. 32.
- Vega, Pedro, J.S., and others (2024). Convolutional neural networks for hydrothermal vents substratum classification: An introspective study. Ecological Informatics, vol. 80, art.102535.
- Versteegh, Emma A., and others (2023). Multiple nutritional strategies of hydrothermal vent shrimp (Rimicaris hybisae) assemblages at the Mid-Cayman Rise. Deep-Sea Research, vol. 192, art. 103915.
- Vrolijk Peter, and others (2021). Using a ladder of seeps with computer decision processes to explore for and evaluate cold seeps on the Costa Rica active margin. Frontiers In Earth Science, vol. 9, art. 601019.
- Waghorn, Kate, and others (2022). Svyatogor Ridge - A Gas Hydrate System Driven by Crustal Scale Processes. World Atlas of Submarine Gas Hydrates in Continental Margins, pp. 247-254.
- Watanabe, Hiromi, and Takehisa Yamakita (2022). Designation of offshore seafloor Natural Environment Conservation Area in the Japanese EEZ. Deep-Sea Life, 18, p. 22. https://www.dosi-project.org/wp-content/uploads/DSL18_final.pdf.
- Workman, Evelyn, and others (2024). Methane emissions from seabed to atmosphere in polar oceans revealed by direct methane flux measurements. Journal of Geophysical Research: Atmospheres, vol. 129, art. e2023JD040632.
- Zeng, Xiang, and others (2020). Genome sequencing of deep-sea hydrothermal vent snails reveals adaptations to extreme environments. GigaScience, vol. 9, No. 12, art. Giaa139.
- Zhou, Yadong, and others (2022). Delineating biogeographic regions in Indian Ocean deep-sea vents and implications for conservation. Diversity and Distributions, vol. 28, No. 12, pp. 2848-2870.