The role of ecosystems in the carbon cycle
Writing team: Steve Widdicombe (coordinating author), Catia Fernades Barbosa, Punyasloke Bhadury, Rafael González-Quirós (lead member), Udhi Eko Hernawan, Natalie Hicks (lead member, part 3), Hiroshi Kitazato, Lisa Levin, Chao Liu, Patrick Martin (lead member, part 4), Ahmed Hussein Mohamed, Alex Poulton (lead member, part 2), Renato Quinones (co-lead member), Olivia Rendon (lead member, part 5) and Sevrine Sailley.
Key points
- Covering approximately 71% of the Earth's surface, the open ocean contributes to approximately half of annual primary production, supporting approximately 80% of global animal biomass.
- The open ocean currently sequesters a similar amount of atmospheric CO2 to that sequestered by the whole terrestrial biosphere, and the ocean has the capacity to store 40 times more CO2 than the atmosphere.
- In coastal systems, coastal blue carbon ecosystems, such as seagrass, mangroves and saltmarshes, are estimated to capture and store more carbon per unit area than terrestrial forests, meaning that, despite covering an area of less than 7 million km2, coastal blue carbon ecosystems are considered globally important areas of carbon capture and storage, which contributes to their high economic and social value, together with other ecosystem services.
- Unvegetated, sea floor sediments on the continental shelf make up approximately 9% of the total marine area but hold the biggest carbon store in shelf systems, and more than 80% of organic carbon is buried in these subtidal sediments.
- The protection and restoration of coastal blue carbon ecosystems and shelf sediment seabed habitats will have significant benefits for both climate mitigation and biodiversity conservation.
- Continental shelf seas are a significant source of carbon (0.95 ± 0.25 Pg C y-1) to the open ocean, although this transport is largely driven by physical and geochemical processes, rather than biological. However, horizontal carbon transport through the migration of animals is probably widespread, but largely unquantified.
- The value of carbon cycling ecosystems has been increasingly recognized, and many finance initiatives exist. However, blue carbon activities receive just 3% of total climate investment globally. This lack of financial support has been a barrier to bringing blue carbon conservation and restoration activities to scale.
- The recognition of the value of coastal blue carbon ecosystems in climate action has increased in recent years. However, at the time of publication, just over half (56%) of Parties to the United Nations Framework Convention on Climate Change have integrated coastal and marine nature-based solutions within new or updated nationally determined contributions as part of mitigation or adaptation measures.
1. Introduction
While the vast majority of the carbon in the ocean is inorganic, and many of the processes that fix, transform and transport this carbon within and through the ocean are driven by physical or chemical processes (see sect. 4, chap. 3), there are also important biological processes that contribute to significant carbon cycling, and these are directly linked to the presence, function and health of marine ecosystems. Given that these same ecosystems also provide vital services to humans, including food provision (see subsect. 5A, chap. 1), understanding and protecting the role of ecosystems in driving the carbon cycle is intrinsically linked to the health (see subsect. 5B, chap. 2) and well-being of billions of people (see subsect. 5B, chap. 3). In this chapter, we will explore the role played by biological processes of key ecosystems in cycling carbon through the two main ocean systems: the open ocean (part 2); and the near shore environment, comprising shelf, coastal, fjordic and estuarine systems (part 3). In part 4 of the present chapter we will describe the transport pathways of carbon between the near shore and the open ocean. Lastly, we will consider the potential values of these ecosystems, and the carbon cycling processes that they provide, in the context of economic and non-economic valuation (part 5).
2. The open ocean
Carbon cycling in marine ecosystems
Accounting for approximately 71% of the Earth's surface, the open ocean contributes to approximately half of Earth's annual primary production (~49–56 Pg C yr-1) Ref 13 Ref 68, despite much lower levels of photosynthetic biomass in the ocean (~0.5-2.4 Pg C) than on land (~450 Pg C) Ref 14 Ref 6. Primary production is dominated by microbial phytoplankton in the open ocean Ref 14 Ref 6, with lifespans ranging from a few days to weeks, leading to an inverted biomass pyramid from microbial primary producers at the base, up to higher trophic levels at the top, and a rapid turnover of photosynthetic biomass in the ocean (see figure I). In the wider context of the global carbon cycle, the marine biosphere represents a relatively small component; while marine biological (organic) carbon pools total approximately 5 Pg C, the ocean contains approximately 38,000 Pg C of dissolved inorganic carbon (DIC) and the atmosphere approximately 800 Pg C (see figure I; Ref 37). However, this relatively small amount of biomass, with significant fluxes between biological pools (see figure I), supports the life and productivity of the global ocean, the largest ecosystem (biome) on the planet.
Figure I Open ocean ecosystem roles in the carbon cycle
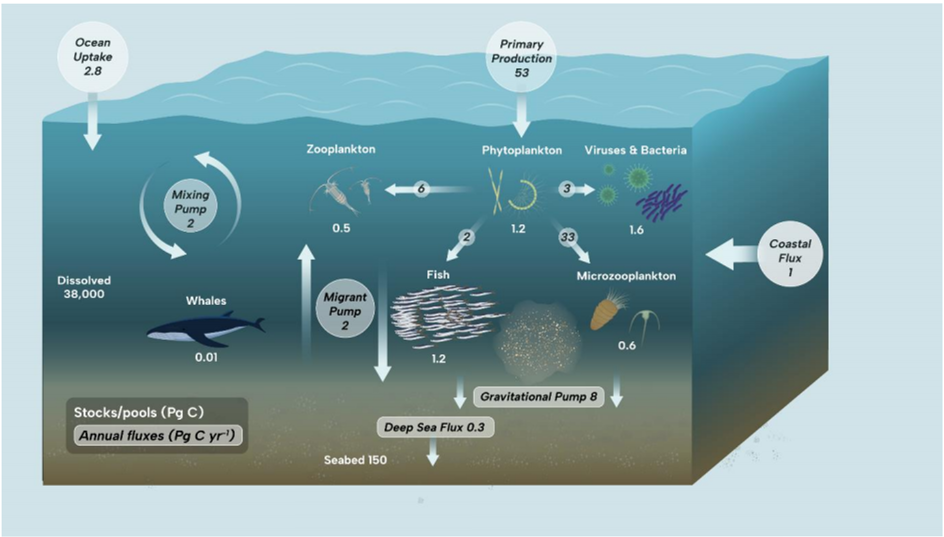
Significant rates of primary production and rapid turnover allow the ocean to support approximately 80% of global animal biomass and approximately half of global protist biomass, which includes significant biomass from pelagic and mesopelagic fish (0.7-1.7 Pg C), large zooplankton (e.g. copepods, krill) (0.3- 0.6 Pg C) and small microzooplankton, including protists such as ciliates and dinoflagellates (0.5-0.7 Pg C) Ref 14 Ref 6 Ref 7 Ref 9 Ref 28. While marine mammal biomass is roughly twice the size of wild terrestrial mammal biomass, it represents a low level of biomass overall (~0.007 Pg C), with baleen whales making up the majority Ref 46. Globally, marine mammal biomass is an order of magnitude smaller than the human population (0.065 Pg C) or domesticated mammal biomass (0.1 Pg C) Ref 6 Ref 46.
Carbon fixed into marine biomass through photosynthesis in the open ocean is rapidly consumed by pelagic organisms, with approximately 62% lost to ingestion and respiration by microzooplankton, approximately 12% directly to larger zooplankton and approximately 2% to 10% to viral lysis and infection Ref 16 Ref 110 Ref 40. Smaller proportions of primary production are lost directly, or indirectly through grazing on herbivorous zooplankton, to fish (<3%; Ref 9) and marine mammals. The remaining proportion of primary production (~18% or ~11 Pg C yr-1) is lost from the upper ocean through the sinking of particulate material (i.e. cells, detritus), or the downward fluxes of dissolved organic material, or through the active migration of pelagic organisms from surface waters to depth Ref 11 Ref 112.
Biological carbon pump(s)
While the open ocean lacks the immense plant biomass of land ecosystems, the ocean currently sequesters a similar amount of anthropogenic atmospheric CO2 (~2.8 Pg C yr-1) to that sequestered by the whole terrestrial biosphere Ref 37, mainly through its physical circulation and geochemical characteristics (Ref 105; see sect. 4, chap. 3), although open ocean ecosystems also play a crucial role Ref 11 Ref 112 Ref 35. Marine photosynthesis draws CO2 from the atmosphere into surface biomass, a portion of which enters the interior ocean through one of several transport pathways (see figure I), driving a gradient in CO2 from the upper ocean into the deep sea through the collective action of ocean ecosystems, termed the biological carbon pump Ref 11 Ref 112. While most of this material is typically broken down and respired by bacteria and zooplankton in the upper 1 km of the ocean, a small proportion of surface production (<2%) does reach the sea floor. So, although the biological carbon pump does not actually contribute much to the net ocean C sink, largely because its vertical fluxes (export) are balanced by the release of absorbed CO2 (outgassing) from upwelled deep waters, it does maintain the long-term store of carbon in deep waters and sediments that would otherwise reside in the atmosphere Ref 35.
The timescale of ocean mixing is proportional to water depth. Some deep waters can be isolated from the atmosphere for up to approximately 1,000 years, while the upper ocean (<0.5 km) may exchange with the atmosphere seasonally, such as during winter mixing. The deeper a sinking particle reaches into the ocean the longer the carbon associated with that particle will be sequestered from the atmosphere. As a global average, particles sinking below 500 m are sequestered for 200 years or longer Ref 11, but this varies geographically depending on regional ocean circulation and mixing Ref 5.
The breakdown and respiration of organic matter supplied to the ocean's interior through the biological carbon pump contributes to the ocean's capacity to store 40 times more CO2 than the atmosphere Ref 37 Ref 35 and fuels deep sea bacteria and fauna at the sea floor. An even smaller proportion (<1%) of surface production ends up sequestered into deep-sea sediments through the physical activity of organisms living at the sea floor. However, the vast area of the deep sea means that this small fraction of surface production makes up a large carbon stock that contributes significantly to C cycling and sequestration on millennial timescales (150-2322 Pg C, depending on whether the upper 5 cm or 1 m is considered; Ref 4) which is similar in size to that stored in terrestrial soils (1700 Pg C) or permafrost (1400 Pg C) Ref 37.
Sinking particulate organic carbon (POC) falls as cells and detrital material, often aggregated into particles termed "marine snow" or packaged by large zooplankton as faecal pellets, and contributes to the "gravitational pump" which is responsible for ~80% of the approximately 11 Pg C annually exported by the biological carbon pump Ref 11 Ref 112. Often these sinking particles still hold nutrients usable by heterotrophs, and it is now recognized that zooplankton often fragment these sinking particles into smaller particles that slow their descent, keeping them in the upper ocean longer Ref 82. This strategy has been termed "microbial gardening" where zooplankton promote bacterial and protist growth on fragmented particles before consuming the enriched microbial biomass Ref 82 which are more nutritious than the original sinking material Ref 18. This highlights the intricate ecological interactions involved in regulating carbon flow into the deep ocean.
Dissolved organic carbon (DOC), varying in form from simple metabolites such as polysaccharides and proteins to complex reworked molecules such as fulvic and humic acids, is an important component of the ocean's dissolved carbon pool Ref 37 with microbial utilization of this pool as a carbon and nutrient source, responsible for breaking down the labile fractions of this material Ref 47 Ref 57 Ref 73. This "microbial carbon pump" recycles carbon and nutrients from the dissolved pool but also changes the nature of the pool to a more refractory one which contributes to long-term carbon sequestration, lasting from hundreds to thousands of years Ref 47 Ref 58 Ref 73. Viral infection also plays a role in the microbial carbon pump, with viral lysis of infected plankton in the upper ocean providing a "viral shunt" retaining carbon near the surface, where it is broken down by microbes Ref 113 Ref 65. Recent research shows that some viral infections do not cause immediate cell lysis. Instead, metabolically inactive infected cells may sink, creating a "viral shuttle" that transports organic material to the deep ocean Ref 66 Ref 99.
The remaining carbon exported through the biological carbon pump is pumped to depth through two distinct mechanisms, one related to the physical mixing of the water column ("mixing pump") and the other related to vertical migration by zooplankton and nekton ("migrant pump") Ref 11 Ref 112. The "mixing pump" involves various oceanographic processes that transport surface water into the ocean's interior, and any dissolved and particulate carbon contained in this water thereby becomes sequestered in the deep ocean. The volume of water, and the depth to which it is transported, varies seasonally and geographically.
Many organisms in the ocean go through daily or seasonal migrations from surface waters into the ocean's interior, either driven by avoidance of visual predators during daylight or to avoid suboptimal growth conditions. These migrations carry carbon to depth, either within the organisms' bodies, to be excreted at depth as faecal pellets or respired over time and released as dissolved CO2. This "migrant pump" transports approximately the same amount of carbon to depth (~2 Pg C yr-1) as the physical mixing of the ocean Ref 11 Ref 112. This behaviourally driven flux of carbon into the deep ocean is extremely important as these migrations are often down to significant depths (>1 km) where the carbon released may be sequestered away from the atmosphere for more than 500 years Ref 11.
Environmental controls, climate change and anthropogenic impacts
While deep remineralization of sinking material is advantageous in terms of sequestering atmospheric CO2 in the ocean's interior, it robs the sun-lit upper ocean of nutrients and leads to widespread nutrient limitation of primary production Ref 91. Processes by which nutrients are returned to the surface, often through stochastic or seasonal mixing, are therefore critical to support continued production. Ocean warming, as a consequence of the ocean absorbing 90% of the total energy generated by anthropogenic forcing of the climate system Ref 21, is likely to alter ocean circulation and mixing (Melet and others, 2022; see sect. 4, chap. 3). While the specific timing and intensity of the changes are unclear, it is likely to result in changes to the physical replenishment of essential nutrients required for primary production, growth and ecosystem health (Ref 54). Current IPCC-class global models vary considerably in their spatial projections of future changes in primary production. Although most exhibit only a modest reduction (~2%) in global marine productivity, there may be strong (>20%) increases (e.g. Arctic) and decreases (e.g. subpolar Atlantic) Ref 114 in specific regions.
Another concern is that reduced ocean mixing will reduce the replenishment of oxygen to deep waters (see sect. 4, chap. 3), essential for heterotrophic activity and higher trophic levels. The respiration of sinking organic matter requires oxygen, so there is a net oxygen consumption (deoxygenation) below the sunlit upper ocean. In shallow areas, or where there is sluggish circulation and high rates of sinking organic material and oxygen consumption, this deoxygenation can lead to hypoxic or anoxic conditions Ref 86, the formation of oxygen minimum zones and avoidance of these areas by aerobic organisms Ref 109. Reductions in ocean mixing due to ocean warming are predicted to lead to the expansion of oxygen minimum zones and the lowering of oxygen concentrations more generally Ref 109 (see sect. 4, chap. 3), which will have a direct impact on ocean biology; for example, limiting the vertical migration of zooplankton Ref 1 and the strength of the migrant pump component of the biological carbon pump Ref 17. Reductions in oxygen availability in the ocean interior may also reduce respiration and remineralization rates, acting as feedback to the biological carbon pump through deeper depth penetration of sinking material (i.e. higher C sequestration) Ref 17.
The ocean's increased uptake of atmospheric CO2, relative to pre-industrial times, is primarily driven by increasing atmospheric CO2 concentrations Ref 105, with the ocean's role in sequestrating anthropogenic CO2 resulting in chemical changes in the ocean Ref 105. Specifically, the dissolution of CO2 and its reaction with seawater to form carbonic acid has dramatically decreased ocean pH (by increasing proton concentrations) and reduced the saturation state of carbonate minerals (calcite, aragonite) in the ocean over the past few decades Ref 71 Ref 56. This "ocean acidification" impacts ocean biology by making the balancing of cell pH (avoiding acidosis) more challenging for marine organisms and increasing the physiological costs of countering the increased dissolution rates of organism-associated biominerals. Ocean acidification impacts the role of marine ecosystems in the carbon cycle at multiple scales (Doney and others, 2020), for example changes in nutrient availability (e.g. Ref 111), growth, grazing and reproductive success Ref 59, calcification, plankton diversity and community composition Ref 116. Despite considerable knowledge of ocean acidification impacts on specific elements of marine ecosystems and the biological carbon pump Ref 116, a holistic understanding of its impact on the future magnitude and efficiency of the different component pumps (gravitational, migrant, mixing) is lacking Ref 94 Ref 125.
3. Shelf, coasts, fjords and estuaries
Moving in from the open ocean, into the shallower areas around coasts, the surface waters continue to fix and cycle carbon through many of the biological processes described in the previous part of the chapter. However, unlike open ocean environments, coastal areas are more complex and varied, often made up of different, interconnected coastal habitats (such as estuaries (see sect. 4, subchap. 5F), mudflats (see sect. 4, subchap. 5B), saltmarshes (see sect. 4, subchap. 5I), seagrass meadows (see sect. 4, subchap. 5G), lagoons and mangroves (see sect. 4, subchap. 5H)), linking the terrestrial zone to the marine through rivers which meet the ocean through deltas, estuaries or fjords (see sect. 4, subchap. 5R). This riverine connection transports organic and inorganic carbon, along with fine particles, into the coastal region, depositing carbon and fine sediment particles (mud) in the nearshore environment Ref 10. The input of terrestrial carbon into the marine environment is reflected in the carbon sequestered in coastal habitats, which is often a mix of terrestrial and marine carbon, with the proportion of terrestrial carbon higher closer to the coast and marine carbon dominating with greater distance from land Ref 124.
In addition, in coastal zones where sufficient light can reach the sea floor, the seabed plays an additional role in carbon fixation Ref 74 Ref 41. Coastal primary production is driven by photosynthesis in plants (see sect. 4, subchap. 4H), macroalgae (see sect. 4, subchap. 4I) or small single celled photosynthetic organisms (microphytobenthos), which inhabit the surface of soft sediment habitats, such as intertidal mudflats Ref 22. Microphytobenthos dominate carbon cycling in unvegetated coastal sediments Ref 119, play a role in pollutant remediation and form the basis of food webs (Hope and others, 2019). However, the complexity of coastal systems means that, currently, considerable uncertainties remain with regard to estimating the actual area covered by these vegetated coastal habitats, along with considerable variability in specific carbon flux estimates, resulting in a tenfold bracket around the estimates of their contribution to organic carbon sequestration, from 0.073 to 0.866 Pg C yr-1 Ref 29.
Coastal blue carbon ecosystems
Vegetated coastal habitats such as salt marshes (see sect. 4, subchap. 5I), seagrass meadows (see sect. 4, subchap. 5G) and mangroves (see sect. 4, subchap. 5H), were first described as "blue carbon" habitats in 2009 Ref 93, and the term is now more generally used for biologically driven carbon fluxes and storage in marine systems that are amenable to management (IPCC, 2021) Ref 76. It is estimated that these three coastal blue carbon ecosystems capture and store more carbon per unit area than terrestrial forests Ref 83, meaning that, despite covering an area of less than 7 million km2 (Duarte 2017), coastal blue carbon ecosystems are considered globally important areas of carbon capture and storage. The plants in these ecosystems capture CO2 directly through photosynthesis (see figure II), and much of the carbon remains within that ecosystem, buried in the soft sediments and root systems around the plants Ref 77 Ref 106. The sediments in these plant-rich ecosystems are key for long-term carbon accumulation and storage, and this permanence (long-term storage of carbon) provides the blue carbon benefit and the potential for climate mitigation Ref 122. However, not all the carbon stored in coastal blue carbon ecosystems is derived from that habitat (autochthonous), and allochthonous carbon (produced outside of that habitat, e.g. terrestrial origin) can make up a significant portion of carbon stocks in sediments Ref 24 Ref 63. This can have implications for blue carbon accounting (see sect. 4). In addition, a substantial contribution to the carbon sequestration potential of coastal blue carbon ecosystems can come from the export of alkalinity that is produced by anaerobic processes (especially denitrification and sulfate reduction) in sediments and exported laterally through physical water movement (Santos and others, 2021).
The original definition of blue carbon has expanded in recent years as scientific knowledge around carbon uptake and storage in marine ecosystems has increased. Unvegetated marine sediments (coastal, shelf and offshore) store significant amounts of carbon owing to their geographical extent Ref 74, despite low accumulation rates compared with coastal zones. The term blue carbon has broadened beyond the original coastal blue carbon ecosystems to include marine sediments (including coastal sediments such as mudflats Ref 12 and continental shelf and offshore sediments Ref 44), which store carbon for climate-relevant (at least 100 years) and geological time periods (>1,000 years) but do not directly capture it like vegetated soft sediment habitats (EMB policy brief on blue carbon, 2023). Shelf sediments (see sect. 4, subchap. 5B) make up ~ 9% of the total marine area Ref 26 but hold the biggest carbon store in shelf systems Ref 74 and more than 80% of organic carbon is buried in subtidal sediments Ref 48 Ref 27. The amount of carbon stored in shelf sediments varies with sediment type, and although finer particles and muddier sediments are associated with higher organic carbon content Ref 10 Ref 74 Ref 26, most of the continental shelf is covered by permeable, sandy sediment, or a mix of mud and sand Ref 27. Sandy sediments hold less organic carbon but a large amount of inorganic carbon, which is generally not considered as part of blue carbon.
Figure II Major carbon pathways and stores in coastal marine ecosystems
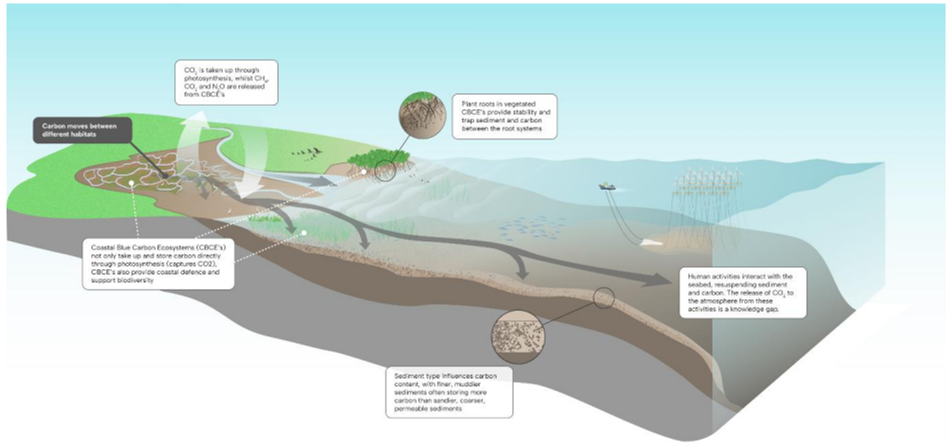
As shown in figure II, carbon is directly captured by plants (as carbon dioxide (CO2)) through photosynthesis in coastal blue carbon ecosystems, while methane (CH4), nitrous oxide (N2O) and CO2 are released by coastal blue carbon ecosystems. Carbon may become stored in coastal blue carbon ecosystems, particularly in the sediment around the root systems, or may flow between the different but connected coastal ecosystems and out onto the shelf (as indicated by the arrow). The largest carbon stores are in the sediments, and carbon stocks vary with sediment type. Human activities (such as fisheries activity or energy infrastructure) that interact with the seabed resuspend sediment and carbon.
Macroalgae and calcifiers
Other organisms play a major role in the coastal biological carbon cycle, such as macroalgae (see sect. 4, subchap. 4I) and calcifying organisms. Macroalgae (such as kelp and seaweed) take up CO2 through photosynthesis and release it at the end of a growing season when much of the algal biomass dies. Since this vegetation is not rooted in sediment, much of this biomass goes through degradation and remineralization (microbial breakdown into CO2). Up to 40% may be released into the coastal and open ocean Ref 67, and a small proportion may be buried in marine sediments Ref 100. Calcifying organisms, such as corals (see sect. 4, subchaps. 5D and 5E) and bivalves (e.g. mussels or oysters), produce solid calcium carbonate in the form of skeletons and shells. While it may seem counter-intuitive, the process of calcification actually emits CO2 (Pernet and others, 2024), and many calcifiers additionally produce CO2 through respiration. However, calcifying organisms do play a vital role in the biological carbon cycle, even though they are not considered to be blue carbon, or a nature-based solution for climate change. They are also important for many other species and provide a number of co-benefits, such as habitat provision, supporting biodiversity and improved water quality (see sect. 4, subchaps. 5D and 5E). Recently, coralline algal beds found in sediments have also been suggested to be blue carbon habitats, because the carbon captured and stored through photosynthesis may exceed the carbon released by calcification Ref 55.
Microbial carbon cycling
As in the open ocean, microbial communities make up a significant proportion of the carbon biomass (see part 2 of the present chapter) and drive sedimentary carbon dynamics on the seabed (balance between sequestered and respired). Organic carbon reaches the seabed and is either remineralized through microbial breakdown or consumption of the carbon, releasing CO2 and nutrients Ref 89 or a small proportion may become buried in the sediment and eventually sequestered. The proportion of carbon buried rather than respired is driven not only by microbial activity, which favours aerobic respiration Ref 45, but also by environmental parameters such as temperature and water depth Ref 26 and presence of macrofauna Ref 89. Generally, warmer waters stimulate more microbial activity, and a greater proportion of carbon is converted to CO2 while less is buried. This has implications for reducing carbon sequestration as ocean temperatures warm. Microbial processing of organic carbon determines the flux of oxygen and carbon dioxide (CO2) at the sediment-water interface (and the sea surface-air interface) and methane in coastal sediments Ref 70.
Carbon is microbially mineralized in the surface oxic layers of the sediment, the depth of which varies with sediment type and season (Hicks and others, 2017), as muddy sediments with smaller grain size have a shallower oxic depth and have higher organic carbon content than sandy sediments Ref 10. Below the oxic layer in sediments, the carbon is less easily broken down by microbial communities owing to the hypoxic or anoxic conditions. However, hypoxic or anoxic microbial breakdown of carbon can lead to the release of other metabolites such as ammonium, hydrogen sulfide or methane, another greenhouse gas Ref 90, although a greater proportion of carbon becomes buried in sediments under low-oxygen conditions. This has implications for warming of coastal and marine ecosystems, leading to an increase in the occurrence, duration and frequency of hypoxia Ref 25.
Coastal microbial carbon cycling in coastal blue carbon ecosystems is influenced by the presence of plant roots, which provide a route for oxygen to penetrate the sediment layers beyond the oxic zone (Ref 117), and microbial communities differ between the coastal blue carbon ecosystem habitats. The origin of the carbon (allochthonous rather than autochthonous) is also important for microbial processing, as marine microbes are less efficient at breaking down land-derived (allochthonous) carbon than marine-derived (autochthonous) carbon Ref 24 Ref 70.
Knowledge gaps and uncertainties
The protection and creation of coastal blue carbon ecosystems are seen as nature-based solutions for marine climate change mitigation Ref 98 and may contribute to nationally determined contributions under the Paris Agreement 2015, for climate mitigation Ref 79. Recent estimates suggest that, with full restoration of coastal blue carbon ecosystems, the total contribution to climate mitigation is approximately 2% globally. In temperate regions, the small geographical area of these ecosystems means that it is unlikely that they will play a significant role in meeting mitigation targets. For small island nations, which have a larger geographical extent of coastal blue carbon ecosystems, their relative contribution may be significantly higher. For example, mangroves are found in more than 100 countries, but more than a fifth of global mangroves are located in Indonesia Ref 78 Ref 43. The huge global variability of carbon stocks in the same habitat makes it difficult to accurately assess carbon sequestration and climate change mitigation potential, and studies are often underrepresented in the global South Ref 121.
Recent research has been focused on different approaches to restoration and conservation of coastal blue carbon ecosystems for climate mitigation, particularly as they are vulnerable to sea level rise in the next few decades. Restoration efforts could include intentional breaching of sea defences or embankments in North-West Europe (called "managed realignment") Ref 15 Ref 92. Restoration approaches for coastal blue carbon ecosystems, such as realignment, provide a nature-based solution for increasing carbon capture and storage, as well as providing coastal protection from floods and storm surges. However, the role of greenhouse gases in carbon cycling, particularly in coastal blue carbon ecosystems, is not yet fully understood Ref 122 (EMB policy brief on blue carbon, 2023), and the climate benefit of salt marshes (natural and restored) is variable Ref 80 Ref 81 Ref 123. Studies have shown that coastal wetlands can be emitters of greenhouse gases such as methane and nitrous oxide Ref 85 Ref 80 Ref 101, although this may be linked to the health and age of the coastal wetland.
Restoration of coastal blue carbon ecosystems for climate mitigation (ability to store carbon) would lead to co-benefits such as coastal protection, increased biodiversity and improved water quality Ref 122. This is important to recognize, because the climate benefits of restoration, or rewilding, efforts are not likely to be immediate, and it will take time for climate mitigation objectives to be met through the transition of other habitats into coastal blue carbon ecosystems (Ref 96) Ref 15 Ref 123. Coastal blue carbon ecosystems will also need to be climate resilient, in the face of warming and rising sea levels Ref 78.
Coastal and shelf soft sediments are a significant repository for carbon, and shelf sediments are often highly productive systems with intensive seabed activity. Despite concerns that these carbon stocks could be remineralized into CO2 through disturbance of the sedimentary carbon stores (Sala and others, 2021), there is limited empirical evidence on the vulnerability of this carbon to anthropogenic activity Ref 32 (Ref 49). This includes trawling, dredging, installation and removal of energy structures and associated infrastructure and deep-sea mining. The uncertainty around the vulnerability of the carbon, and limited knowledge on carbon accumulation rates and burial, makes it challenging to effectively manage the seabed.
4. Exchange between coastal and open ocean systems
In previous parts of the chapter the open ocean and the coastal environment have been considered separately. However, these two systems are tightly coupled when it comes to carbon transport, with continental shelf seas being a source of 0.95 ± 0.25 Pg C yr-1 to the open ocean Ref 102. This flux consists chiefly of dissolved inorganic carbon, dissolved organic carbon and particulate organic carbon (DIC, DOC and POC, respectively) and is mainly driven by physical processes. The net transport of DIC is largely driven by enhanced physico-chemical dissolution of atmospheric CO2 into shelf waters. Shelf seas also typically have higher concentrations of DOC and POC than the open ocean, because of high biological productivity and inputs of DOC and POC from land. Moreover, movement of nektonic animals to and from shelf seas contributes a carbon flux, although this remains very poorly quantified. Quantifying these carbon fluxes is challenging, in part because of the high spatial and temporal variability of shelf seas.
The purely physico-chemical mechanism of shelf-to-ocean DIC flux is mainly driven by winter-time cooling of shelf waters at temperate and higher latitudes, which lowers the partial pressure of CO2 (pCO2) in the water and results in uptake of atmospheric CO2. This cold, dense shelf water can then flow off the shelf and below the oceanic thermocline, a process called the "continental shelf pump" Ref 118 Ref 23. Conversely, in tropical shelf seas the opposite can occur: cooler oceanic water can warm up on the shelf, leading to an increase in shelf sea pCO2 and promoting CO2 flux to the atmosphere Ref 69.
Biological production contributes to the continental shelf pump by converting CO2 into POC and DOC. However, to become part of the continental shelf pump this POC and DOC must either be transported below the open-ocean thermocline or be transformed into refractory DOC, so that the carbon is not remineralized back into CO2 and exchanged with the atmosphere over short timescales Ref 72 Ref 50. However, most of the POC and DOC produced in shelf seas is remineralized before it can become part of the continental shelf pump, and net biological production rates are not correlated with the strength of the CO2 sink when comparing globally across shelf seas Ref 69. Moreover, there can be seasonal and even inter-annual lags between biological production and physical transport, such that shelf sea carbon budgets are not necessarily balanced over a year Ref 20 Ref 53. The production of biologically refractory DOC, which can accumulate and be exported off the shelf during episodic flushing events, mass transportation of both DOC and POC from shelf to the deep sea by tectonic triggers such as big earthquakes and tsunamis Ref 60, is therefore important for the continental shelf pump; since this carbon is refractory it does not matter whether this material is exported above or below the thermocline Ref 20 Ref 53. Part of the POC and DOC that is transported from shelf seas to the open ocean is terrestrial in origin. The fate of this terrestrial carbon remains poorly quantified, but approximately 50% appears to be transported off shelf seas (Ref 69), including below the permanent thermocline Ref 84. Turbidity currents are an important vector transferring shelf sea sedimentary POC below the oceanic thermocline Ref 103 Ref 42 Ref 115. A distinct increase in coastal to upper slope temperatures gives rise to unstable conditions in the organic rich sediments found under the oxygen minimum zone. The high rate of organic carbon deposition can introduce slope failures by the dissolution of gas hydrates. These non-tectonic slope failures may be an important starting point for the lateral transportation of organic carbons to the deep ocean in the form of turbidites Ref 64.
Some organic matter enters the open ocean above the permanent thermocline. Unless it is in the form of refractory DOC, this carbon is not sequestered from the atmosphere, but it can be recycled and fuel primary production off the shelf, and also contribute to the biological carbon pump in the open ocean Ref 38. This contribution is partly directly by POC and DOC originating from the shelf, and partly by POC and DOC produced from recycling of organic matter that originated from the shelf.
Horizontal carbon transport through animals is probably widespread but largely unquantified, unlike the vertical flux from animal migrations (see part 2 of the present chapter). For example, fish contribute 16% ± 13% of downward POC flux out of the euphotic zone, but the horizontal carbon flux from fish migrations is unknown Ref 104. Physical transport of zooplankton across the shelf break can be significant Ref 62 and is even part of the ontogenetic migrations of some commercially important species such as rock lobsters Ref 97. Horizontal fluxes have been estimated in only a few instances. For example, the spawning migration of the squid Illex argentinus on the Patagonian Shelf transfers approximately 0.09 Gg C yr-1 (0.09x10-6 Pg yr-1) to the shelf slope Ref 3, using biomass-to-carbon conversions from Ref 52). This is approximately 0.7% of the Patagonian Shelf's net biological production Ref 69, although it is partly balanced by on-shelf migration of other squid species. Horizontal carbon transport via nektonic animals is likely to be impacted by fishing practices and potentially by climate-induced changes in seasonal and ontogenetic migration patterns.
5. The value of carbon cycling ecosystems
Ocean natural capital accounting (encompasses the systematic measurement and reporting of the status or "health" of ocean natural capital, as well as the contributions these assets make to society and the blue economy. Through monetary quantification, the ocean natural capital accounting enables decision makers to make trade-offs and assess impacts associated with various development scenarios and policies for ocean environmental sustainability and human well-being. Marine and coastal ecosystems form natural capital stocks that provide numerous benefits - such as food, climate regulation and air quality control - collectively known as ecosystem services. The value of coastal blue carbon ecosystems can be understood as the total economic value of conserving, restoring and sustainably managing marine and coastal ecosystems, while the concept of total economic value comprises the use and non-use values (Zarate- Barrera and Maldonado, 2015). The valuation of the ecosystem services provided by marine and coastal ecosystems clearly evidences the links between humans and the oceans, informs policymakers for improved ocean management/planning and advance policies and motivates the investment in ocean conservation.
Recently, a country-level blue carbon wealth assessment showed that coastal blue carbon ecosystems have an average value of $190.67±30 billion. Australia, Indonesia and Cuba were the three countries that contributed the most benefits from blue carbon wealth and India, China and the United States are the largest beneficiaries Ref 8. Of the different types of coastal blue carbon ecosystems, the most extensively studied are mangroves (see sect. 4, subchap. 5H) and salt marshes (tidal marshes). For instance, all the ecosystem services provided by all mangroves worldwide have been valued at $69.9 billion Ref 107. Regulation services of various coastal wetlands (see sect. 4, subchap. 5I) have an average value of $369.7 and a total economic value of $3,184 ha-1 yr-1. Mangroves have an average of $806 ha-1 yr-1 for provision services, $1,446 ha-1 yr-1 for regulation services, $113 ha-1 yr-1 for support services, $1,721 ha-1 yr-1 for cultural services, and a total economic value of $6,553 ha-1 yr-1 Ref 75. Seagrass meadows (see sect. 4, subchap. 5G) are less studied but have an average $3,226.5 ha-1 yr-1 for provision and $871.7 ha-1 yr-1 for regulation Ref 75. A study examined the ecological and economic potential of three key ecosystem services - fishery production, nutrient cycling, and carbon removal - provided by six kelp genera (Ecklonia, Laminaria, Lessonia, Macrocystis, Nereocystis and Saccharina). The results show that the potential value of each genus was between $64,400 and $147,100 ha-1 yr-1 Ref 30. There are still gaps in valuation of blue carbon ecosystems. For example, Arkema and others (2024) found that only a third of papers quantify societal benefits of coastal ecosystems within marine protected areas (MPAs), and these tend to focus mainly on corals and mangrove ecosystems.
The value of carbon cycling ecosystems has been increasingly recognized and many finance initiatives exist i.e. voluntary and compliance carbon markets, public funding, philanthropic and other alternatives, such as payment for ecosystem services, ocean natural capital accounting and blue bonds. Blue Carbon activities are one type of climate mitigation solution, which currently receives just 3% of total climate investment globally. This lack of financial support has been a barrier to bringing Blue Carbon conservation and restoration activities to scale.
The recognition that marine ecosystems, other than coastal blue carbon ecosystems, also have the power to fix and transport large quantities of carbon has increased interest in using them as marine CO2 removal measures (see subsect. 5A, chap. 10). It should be noted that great uncertainty remains regarding the efficacy, scalability or financial viability of many of these proposed nature-based marine CO2 removal approaches. For example, the estimated costs for ocean iron fertilization vary widely, with best-case values of $7 per net ton carbon captured, compared with worst-case values of $1,500 per net ton carbon captured depending on oceanographic factors. Ocean iron fertilization aerial delivery ranges between $21 and $2,033 per net ton carbon captured inclusive of verification costs; up to 30%-40% more cost-effective than ship-based ocean iron fertilization Ref 31.
While seaweed aquaculture is not considered as either blue carbon or geoengineering (see subsect. 5A, chap. 10), the additional value of such activities to carbon storage, over and above the commercial value of the seaweed product, has been noted. The levelized unit costs (LCOC) of CO2eq sequestration (LCOC; $tCO2eq-1) for kelp aquaculture at baseline were estimated at $17,048 tCO2eq-1, with only a 39% true sequestration "additionality" rate. However, by process improvement and decarbonization of production supply chains LCOC were reduced to $1,257 tCO2eq-1 and the additionality rate increased to 91% (Ref 19). Froehlich and others, (2019) analysed global production data and determined that the cost of producing carbon credits from macroalgae ranged from $71 to $27,222 tCO2eq-1.
More generally, the recognition of the value of coastal blue carbon ecosystems in climate action has increased in recent years. However, while a total of 81% (158) of Parties to the Paris Agreement have included an adaptation component in their nationally determined contributions, only 31% (49) identified ocean ecosystems as a priority sector for adaptation. Of the 21% of Parties that included information on ocean or blue carbon as a priority sector for reducing greenhouse gas emissions, 71% mentioned specific ocean-based mitigation measures Ref 120.
References
- Abedi, E., J. Seyfabadi, A. Saleh, A. Sari (2022). Mesozooplankton community in near-hypoxic and hypoxic layers of the Persian Gulf and Gulf of Oman. Marine Pollution Bulletin, 184, 114146. https://doi.org/10.1016/j.marpolbul.2022.114146.
- Arkema, K.K., L. Field, L.K. Nelson, N.C. Ban, C. Gunn, S.E. Lester (2024). Advancing the design and management of marine protected areas by quantifying the benefits of coastal ecosystems for communities. One Earth, 7: 989-1006. https://doi.org/10.1016/j.oneear.2024.04.019.
- Arkhipkin, A.I. (2013). Squid as nutrient vectors linking Southwest Atlantic marine ecosystems. Deep Sea Research Part II: Topical Studies in Oceanography, 95, 7-20. https://doi.org/10.1016/j.dsr2.2012.07.003.
- Atwood, T.B., Witt, A., Mayorga, J., Hammill, E., Sala, E. (2020). Global patterns in marine sediment carbon stocks. Front. Mar. Sci., 7. https://doi.org/10.3389/fmars.2020.00165.
- Baker, C.A., Martin, A.P., Yool, A., and Popova, E. (2024). Biological Carbon Pump Sequestration Efficiency in the North Atlantic: A Leaky or a Long-Term Sink? Global Biogeochemical Cycles, 36, e2021GB007286. https://doi.org/10.1029/2021GB007286.
- Bar-On, Y., Phillips, R., Milo, R. (2018). The biomass on Earth. PNAS, 115(25),6505-6511. https://doi.org/10.1073/pnas.1711842115.
- Bar-On, Y., Milo, R. (2019). The biomass composition of the oceans: A blueprint of our blue planet. Cell, 179, 1451-1454. https://doi.org/10.1016/j.cell.2019.11.018.
- Bertram, C., Quaas, M., Reusch, T.B.H., and others (2021). The blue carbon wealth of nations. Nat. Clim. Chang., 11, 704-709. https://doi.org/10.1038/s41558-021-01089-4
- Bianchi, D., Carozza, D.A., Galbraith, E.D., and others (2021). Estimating global biomass and biogeochemical cycling of marine fish with and without fishing. Science Advances, 7(41). https://doi.org/10.1126/sciadv.abd7554.
- Bianchi, T.S., Mayer, L.M., Amaral, J.H.F., Arndt, S., Galy, V., Kemp, D.B., Kuehl, S.A., Murray, N.J., Regnier, P. (2024). Anthropogenic impacts on mud and organic carbon cycling. Nature Geoscience, 17(4), 287-297. https://doi.org/10.1038/s41561-024-01405-5.
- Boyd, P.W., Claustre, H., Levy, M., Siegel, D.A., Weber, T. (2019). Multi-faceted particle pumps drive carbon sequestration in the ocean. Nature, 568, 327-335. https://doi.org/10.1038/s41586-019-1098-2.
- Brown, D.R., Marotta, H., Peixoto, R.B., Enrich-Prast, A., Barroso, G.C., Soares, M.L.G., Machado, W., Pérez, A., Smoak, J.M., Sanders, L.M., Conrad, S., Sippo, J.Z., Santos, I.R., Maher, D.T., and Sanders, C.J. (2021). Hypersaline tidal flats as important 'blue carbon' systems: a case study from three ecosystems. Biogeosciences, 18(8), 2527-2538. https://doi.org/10.5194/bg-18-2527-2021.
- Buitenhuis, E.T., Hashioka, T., Le Quere, C.L. (2013a). Combined constraints on global ocean primary production using observations and models. Global Biogeochemical Cycles, 27, 847-858. https://doi.org/10.1002/gbc.20074.
- Buitenhuis, E.T., Vogt, M., Moriarty, R., and others (2013b). MAREDAT: towards a world atlas of MARine Ecosystem DATa. Earth System Science Data, 5(2), 227-239. https://doi.org/10.5194/essd-5-227-2013.
- Burden, A., Garbutt, R.A., Evans, C.D., Jones, D.L. and Cooper, D.M. (2013). Carbon sequestration and biogeochemical cycling in a saltmarsh subject to coastal managed realignment. Estuarine, Coastal and Shelf Science, 120, pp. 12-20. https://doi.org/10.1016/j.ecss.2013.01.014.
- Calbert, A. (2001). Mesozooplankton grazing effect on primary production: A global comparative analysis in marine ecosystems. Limnology and Oceanography, 46(7), 1824-1830. https://doi.org/10.4319/lo.2001.46.7.1824.
- Cavan, E.L., Trimmer, M., Shelley, F., Sanders, R. (2017). Remineralization of particulate organic carbon in an ocean oxygen minimum zone. Nature Communications, 8, 14847. https://doi.org/10.1038/ncomms14847.
- Cavan, E.L., Kawaguchi, S., Boyd, P.W. (2020). Implications for the mesopelagic microbial gardening hypothesis as determined by experimental fragmentation of Antarctic krill fecal pellets. Ecology and Evolution, 11(2), 1023-1036. https://doi.org/10.1002/ece3.7119.
- Coleman S., Dewhurst T., Fredriksson D.W., St. Gelais A.T., Cole K.L., MacNicoll M., Laufer E., Brady D.C. (2022). Quantifying baseline costs and cataloging potential optimization strategies for kelp aquaculture carbon dioxide removal. Frontiers of Marine Science (Sec. Marine Fisheries, Aquaculture and Living Resources), 9. https://doi.org/10.3389/fmars.2022.966304.
- Chaichana, S., Jickells, T., and Johnson, M. (2019). Interannual variability in the summer dissolved organic matter inventory of the North Sea: Implications for the continental shelf pump. Biogeosciences, 16(5), 1073-1096. https://doi.org/10.5194/bg-16-1073-2019.
- Cheng, L., von Schukmann, K., Abraham, J.P., and others (2022). Past and future ocean warming. Nature Reviews Earth and Environment. 3, 776-794. https://doi.org/10.1038/s43017-022-00345-1.
- Christianen, M.J., Middelburg, J.J., Holthuijsen, S.J., Jouta, J., Compton, T.J., van der Heide, T., Piersma, T., Sinninghe Damsté, J.S., van der Veer, H.W., Schouten, S., and Olff, H. (2017). Benthic primary producers are key to sustain the Wadden Sea food web: stable carbon isotope analysis at landscape scale. Ecology, 98(6), pp. 1498-1512.
- Cossarini, G., Querin, S., and Solidoro, C. (2015). The continental shelf carbon pump in the northern Adriatic Sea (Mediterranean Sea): Influence of wintertime variability. Ecological Modelling, 314, 118- 134. https://doi.org/10.1016/j.ecolmodel.2015.07.024.
- Cragg, S.M., Friess, D.A., Gillis, L.G., Trevathan-Tackett, S.M., Terrett, O.M., Watts, J.E., Distel, D.L., and Dupree, P. (2020). Vascular plants are globally significant contributors to marine carbon fluxes and sinks. Annual Review of Marine Science, 12(1), pp. 469-497. https://doi.org/10.1146/annurev-marine- 010318-095333.
- Diaz, R.J., and Rosenberg, R. (2008). Spreading dead zones and consequences for marine ecosystems. Science, 321(5891), pp. 926-929.
- Diesing, M., Kröger, S., Parker, R., Jenkins, C., Mason, C, Weston, K. (2017). Predicting the standing stock of organic carbon in surface sediments of the North-West European continental shelf. Biogeochemistry, 135, pp. 183-200
- Diesing, M., Thorsnes, T., and Bjarnadóttir, L.R. (2021). Organic carbon densities and accumulation rates in surface sediments of the North Sea and Skagerrak. Biogeosciences, 18(6), 2139-2160.
- Drago, L., Panaiotis, T., Irisson, J-O., and others (2022). Global distribution of zooplankton biomas estimated by in situ imaging and machine learning. Front. Mar. Sci., 9. https://doi.org/10.3389/fmars.2022.894372.
- Duarte, C.M. (2017). Reviews and syntheses: Hidden forests, the role of vegetated coastal habitats in the ocean carbon budget. Biogeosciences, 14(2), 301-310.
- Eger, A.M., Marzinelli, E.M., Beas-Luna, R., and others (2023). The value of ecosystem services in global marine kelp forests. Nat Commun, 14, 1894. https://doi.org/10.1038/s41467-023-37385-0.
- Emerson, D., L.E. Sofen, A.B. Michaud, S.D. Archer, B.S. Twining (2024). A Cost Model for Ocean Iron Fertilization as a Means of Carbon Dioxide Removal That Compares Ship- and Aerial-Based Delivery, and Estimates Verification Costs. Earth's Future, 12, e2023EF003732. https://doi.org/10.1029/2023EF003732.
- Epstein, G., Middelburg, J.J., Hawkins, J.P., Norris, C.R. and Roberts, C.M. (2022). The impact of mobile demersal fishing on carbon storage in seabed sediments. Global Change Biology, 28(9), pp. 2875-2894.
- European Marine Board (2023). Blue Carbon: Challenges and opportunities to mitigate the climate and biodiversity crises. EMB Policy Brief No. 11. doi: https://doi.org/10.5281/zenodo.8314215. ISSN: 0778- 3590 ISBN: 9789464206203.
- Faran, T.S., and L. Olsson (2018). Geoengineering: neither economical, nor ethical-a risk-reward nexus analysis of carbon dioxide removal. Int Environ Agreements, 18, 63-77.
- Frenger, I., Landolfi, A., Kvale, K., and others (2024). Misconceptions of the marine biological carbon pump in a changing climate: Thinking outside the 'export' box. Global Change Biology, 30(1), e17124. https://doi.org/10.1111/gcb.17124.
- First Blue Carbon Conservation Methodology Expected to Scale Up Finance for Coastal Restoration & Conservation Activities (2020). https://verra.org/first-blue-carbon-conservation-methodology-expected-to-scale-up-finance-for-coastal-restoration-conservation-activities/.
- Friedlingstein, P., O'Sullivan, M., Jones, M.W., and others (2023). Global Carbon Budget 2023. Earth System Science Data, 15, 5301-5369. https://doi.org/10.5194/essd-15-5301-2023.
- Frischknecht, M., Münnich, M., and Gruber, N. (2018). Origin, Transformation, and Fate: The Three-Dimensional Biological Pump in the California Current System. Journal of Geophysical Research: Oceans, 123(11), 7939-7962. https://doi.org/10.1029/2018JC013934.
- Froehlich, H.E., Afflerbach, J.C., Frazier, M., Halpern, B.S. (2019). Blue Growth Potential to Mitigate Climate Change through Seaweed Offsetting. Current Biology, 29, 3087-3093. https://doi.org/10.1016/j.cub.2019.07.041.
- Gao, Y., Lu, Y., Dungait, J.A.J., and others (2022). The 'regulator' function of viruses on ecosystem carbon cycling in the Anthropocene. Front. Public. Health, 10. https://doi.org/10.3389/fpubh.2022.858615.
- Gattuso, J.P., Gentili, B., Duarte, C.M., Kleypas, J.A., Middelburg, J.J., and Antoine, D. (2006). Light availability in the coastal ocean: Impact on the distribution of benthic photosynthetic organisms and their contribution to primary production. Biogeosciences, 3(4), 489-513. https://doi.org/10.5194/bg-3-489-2006.
- Gibbs, M., Leduc, D., Nodder, S.D., Kingston, A., Swales, A., Rowden, A.A., Mountjoy, J., Olsen, G., Ovenden, R., Brown, J., Bury, S., and Graham, B. (2020). Novel Application of a Compound-Specific Stable Isotope (CSSI) Tracking Technique Demonstrates Connectivity Between Terrestrial and Deep-Sea Ecosystems via Submarine Canyons. Frontiers in Marine Science, 7. https://doi.org/10.3389/fmars.2020.00608.
- Giri, C., Ochieng, E., Tieszen, L.L., Zhu, Z., Singh, A., Loveland, T., Masek, J. and Duke, N. (2011). Status and distribution of mangrove forests of the world using earth observation satellite data. Global ecology and biogeography, 20(1), pp. 154-159.
- Graves, C.A., Benson, L., Aldridge, J., Austin, W.E.N., Dal Molin, F., Fonseca, V.G., Hicks, N., Hynes, C., Kröger, S., Lamb, P.D., Mason, C., Powell, C., Smeaton, C., Wexler, S.K., Woulds, C., and Parker, R. (2022). Sedimentary carbon on the continental shelf: Emerging capabilities and research priorities for Blue Carbon. Frontiers in Marine Science, 9. Available at https://www.frontiersin.org/articles/10.3389/fmars.2022.926215.
- Glud (2008). Oxygen dynamics of marine sediments, Marine Biology Research, 4: 4, 243-289, DOI: 10.1080/17451000801888726.
- Greenspoon, L., Krieger, E., Sender, R., Milo, R. (2023). The global biomass of wild mammals. PNAS, 120 (10), e2204862120. https://doi.org/10.1073/pnas.2204892120.
- Hansell, D.A. (2013). Recalcitrant Dissolved Organic Carbon Fractions. Annual Review of Marine Science, 5(1), 421-445. https://doi.org/10.1146/annurev-marine-120710-100757.
- Hedges, J.I., and Keil, R.G. (1995). Sedimentary organic matter preservation: an assessment and speculative synthesis. Marine Chemistry, 49 (2-3), 81-115.
- Hicks, N., Ubbara, G.R., Silburn, B., Smith, H.E., Kröger, S., Parker, E.R., Sivyer, D., Kitidis, V., Hiddink, J.G., van de Velde, S.J., McConnaughey, R.A., De Borger, E., Tiano, J., Kaiser, M.J., Sweetman, A.K., and Sciberras, M. (2023). Quantifying the carbon benefits of ending bottom trawling. Nature, 617(7960), pp. E1-E2. https://doi.org/10.1038/s41586-023-06014-7.
- Holt, J., Wakelin, S., and Huthnance, J. (2009). Down-welling circulation of the northwest European continental shelf: A driving mechanism for the continental shelf carbon pump. Geophysical Research Letters, 36(14). https://doi.org/10.1029/2009GL038997.
- Hope, J.A., Paterson, D.M., and Thrush, S.F. (2020). The role of microphytobenthos in soft-sediment ecological networks and their contribution to the delivery of multiple ecosystem services. Journal of Ecology, 108(3), pp. 815-830. https://doi.org/10.1111/1365-2745.13322.
- Hoving, H.J.T., Bush, S.L., Haddock, S.H.D., and Robison, B.H. (2017). Bathyal feasting: Post-spawning squid as a source of carbon for deep-sea benthic communities. Proceedings of the Royal Society B: Biological Sciences, 284(1869), 20172096. https://doi.org/10.1098/rspb.2017.2096.
- Humphreys, M.P., Achterberg, E.P., Hopkins, J.E., Chowdhury, M.Z.H., Griffiths, A.M., Hartman, S.E., Hull, T., Smilenova, A., Wihsgott, J.U., Woodward, E.M.S., and Moore, C.M. (2019). Mechanisms for a nutrient-conserving carbon pump in a seasonally stratified, temperate continental shelf sea. Progress in Oceanography, 177, 101961. https://doi.org/10.1016/j.pocean.2018.05.001.
- Hutchins, D.A., Tagliabue, A. (2024). Feedbacks between phytoplankton and nutrient cycles in a warming ocean. Nature Geoscience, 17, 495-502. https://doi.org/10.1038/s41561-024-01451-w.
- James, K., Macreadie, P.I., Burdett, H.L., Davies, I., and Kamenos, N.A. (2024). It's time to broaden what we consider a 'blue carbon ecosystem'. Global Change Biology, 30, e17261. https://doi.org/10.1111/gcb.17261.
- Jiang, L-O., Carter, B.R., Feely, R.A., Lauvset, S.K., Olsen, A. (2019). Surface ocean pH and buffer capacity: past, present, and future. Scientific Reports, 9, 18624. https://doi.org/10.1038/s41598-019-55039-4.
- Jiao, N., Robinson, C., Azam, F., Thomas, H., Baltar, F., Dang, H., Hardman-Mountford, N.J., Johnson, M., Kirchman, D.L., Koch, B.P., Legendre, L., Li, C., Liu, J., Luo, T., Luo, Y.-W., Mitra, A., Romanou, A., Tang, K., Wang, X., and Zhang, R. (2014). Mechanisms of microbial carbon sequestration in the ocean - future research directions. Biogeosciences, 11(19), 5285-5306. https://doi.org/10.5194/bg-11-5285-2014.
- Jiao, N., Luo, T., Chen, Q., Zhao, Z., Xiao, X., Liu, J., Jian, Z., Xie, S., Thomas, H., Herndl, G.J., Benner, R., Gonsior, M., Chen, F., Cai, W.-J., and Robinson, C. (2024). The microbial carbon pump and climate change. Nature Reviews Microbiology, 22(7), 408-419. https://doi.org/10.1038/s41579-024-01018-0.
- Keil, K.E., Klinger, T., Keister, J.E., McLaskey, A.K. (2021). Comparative sensitivities of zooplankton to ocean acidification conditions in experimental and natural settings. Frontiers in Marine Science, 8. https://doi.org/10.3389/fmars.2021.613778.
- Kioka, A., T. Schwestermann, J. Moernaut, K. Ikehara, T. Kanamatsu, C.M. McHugh, C. dos Santos Ferreira, G. Wiemer, N. Haghipour, A.J. Kopf, T.I. Eglinton, M. Strasser (2019). Megathrust earthquake drives drastic organic carbon supply to the hadal trench, Scientific Reports, 9: 1553. https://doi.org/10.1038/s41598-019-38834-x.
- Kowalczyk, K.A., T. Amann, J. Strefler, M.E. Vorrath, J. Hartmann, S. De Marco, P. Renforth, S. Foteinis, E. Kriegler (2024). Marine carbon dioxide removal by alkalinization should no longer be overlooked. Environmental Research Letters, 19: 074033. https://doi.org/10.1088/1748-9326/ad5192.
- Keister, J.E., Peterson, W.T., and Pierce, S.D. (2009). Zooplankton distribution and cross-shelf transfer of carbon in an area of complex mesoscale circulation in the northern California Current. Deep Sea Research Part I: Oceanographic Research Papers, 56(2), 212-231. https://doi.org/10.1016/j.dsr.2008.09.004.
- Kharbush, J.J., Close, H.G., Van Mooy, B.A., Arnosti, C., Smittenberg, R.H., Le Moigne, F.A., Mollenhauer, G., Scholz-Böttcher, B., Obreht, I., Koch, B.P., and Becker, K.W. (2020) Particulate organic carbon deconstructed: molecular and chemical composition of particulate organic carbon in the ocean. Frontiers in Marine Science, 7, p. 518 https://doi.org/10.3389/fmars.2020.00518.
- Kitazato, H. (2024). Global warming may accelerate submarine landslides in the ocean - Possible disaster chain reactions -. Progress in Landslide Research and Technology (Progress-LRT), ICL Open Access Book Series, vol. 3, No. 1, 83-91. https://doi.org/10.1007/978-3-031-55120-8_4.
- Kranzler, C.F., Krause, J.W., Brzezinski, M.A., and others (2019). Silicon limitation facilitates virus infection and mortality of marine diatoms. Nature Microbiology, 4, 1790-1797. https://doi.org/10.1038/s41564-019-0502-x.
- Kranzler, C.F., Brzezinski, M.A., Cohen, N.R., and others (2021). Impaired viral infection and reduced mortality of diatoms in iron-limited oceanic regions. Nature Geoscience, 14, 231-237. https://doi.org/10.1038/s41561-021-00711-6.
- Krause-Jensen, Dorte, and Carlos M. Duarte (2016). Substantial role of macroalgae in marine carbon sequestration. Nature Geoscience, 9.10, 737-742.
- Kulk, G., Platt, T., Dingle, J. (2021). Primary Production, an Index of Climate Change in the Ocean: Satellite Based Estimates over Two Decades. Remote Sensing, 12(5), 826. https://doi.org/10.3390/rs12050826.
- Lacroix, F., Ilyina, T., Laruelle, G.G., and Regnier, P. (2021). Reconstructing the Preindustrial Coastal Carbon Cycle Through a Global Ocean Circulation Model: Was the Global Continental Shelf Already Both Autotrophic and a CO2 Sink? Global Biogeochemical Cycles, 35(2), e2020GB006603. https://doi.org/10.1029/2020GB006603.
- LaRowe, D.E., Arndt, S., Bradley, J.A., Estes, E.R., Hoarfrost, A., Lang, S.Q., Lloyd, K.G., Mahmoudi, N., Orsi, W.D., Walter, S.S., and Steen, A.D. (2020) The fate of organic carbon in marine sediments-New insights from recent data and analysis. Earth-Science Reviews, 204, p. 103146. https://doi.org/10.1016/j.earscirev.2020.103146.
- Lauvset, S.K., Gruber, N., Landschutzler, P., Olson, A., Tjiputra, J. (2015). Trends and drivers in global surface ocean pH over the last 3 decades. Biogeosciences, 12, 1285-1298. https://doi.org/10.5194/bg-12-1285-2015.
- Lee, K., Kim, J.-M., Lee, G.-S., Lee, E., Jeong, J.-Y., Lee, J., and Han, I.-S. (2022). Persistent Continental Shelf Carbon Sink at the Ieodo Ocean Research Station in the Northern East China Sea. Frontiers in Marine Science, 9. https://doi.org/10.3389/fmars.2022.919249.
- Legendre, L., Rivkin, R.B., Weinbauer, M.G., Guidi, L., Uitz, J. (2015). The microbial carbon pump concept: Potential biogeochemical significance in the globally changing ocean. Progress in Oceanography, 134, 432-450. https://doi.org/10.1016/j.pocean.2015.01.008.
- Legge, O., and others (2020). Carbon on the northwest European shelf: Contemporary budget and future influences. Frontiers in Marine Science, 7, 143. https://doi.org/10.3389/fmars.2020.00143.
- Lopez-Rivas, J.D., J.C. Cardenas (2024). What is the economic value of coastal and marine ecosystem services? A systematic literature review. Marine Policy, 161: 106033. https://doi.org/10.1016/j.marpol.2024.106033.
- Lovelock, C.E. and Duarte, C.M. (2019) Dimensions of Blue Carbon and emerging perspectives. Biology Letters, 15(3), 20180781. Available at https://doi.org/10.1098/rsbl.2018.0781.
- Macreadie, P.I., York, P.H., Sherman, C.D.H. (2014). Resilience of Zostera muelleri seagrass to small scale disturbances: the relative importance of asexual verses sexual recovery. Ecology and Evolution 4(4), 450-461. https://doi.org/10.1002/ece3.933.
- Macreadie, P.I., Anton, A., Raven, J.A., and others (2019). The future of Blue Carbon science. Nat Commun, 10, 3998. https://doi.org/10.1038/s41467-019-11693-w.
- Macreadie, P.I., Costa, M.D., Atwood, T.B., Friess, D.A., Kelleway, J.J., Kennedy, H., Lovelock, C.E., Serrano, O., and Duarte, C.M. (2021). Blue carbon as a natural climate solution. Nature Reviews Earth & Environment, 2(12), pp. 826-839.
- Mason, V.G., Burden, A., Epstein, G., Jupe, L.L., Wood, K.A. and Skov, M.W. (2023). Blue carbon benefits from global saltmarsh restoration. Global Change Biology, 29(23), pp. 6517-6545. https://doi.org/10.1111/gcb.16943.
- Mason, V.G., A. Burden, G. Epstein, L.L. Jupe, K.A. Wood, M.W. Skov. (2024). Navigating research challenges to estimate blue carbon benefits from saltmarsh restoration. Global Change Biology, 30: e17526. https://doi.org/10.1111/gcb.17526.
- Mayor, D.J., Sanders, R., Giering, S.L.C., Anderson, T.R. (2014). Microbial gardening in the ocean's twilight zone: Detritivorous metazoans benefit from fragmenting, rather than ingesting, sinking detritus. BioEssays, 36(12), 1132-1137. https://doi.org/10.1002/bies.201400100.
- Mcleod, E., Chmura, G.L., Bouillon, S., Salm, R., Björk, M., Duarte, C.M., Lovelock, C.E., Schlesinger, W.H. and Silliman, B.R. (2011). A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Frontiers in Ecology and the Environment, 9(10), pp. 552-560. https://doi.org/10.1890/110004.
- Medeiros, P.M., Seidel, M., Niggemann, J., Spencer, R.G.M., Hernes, P.J., Yager, P.L., Miller, W.L., Dittmar, T., and Hansell, D.A. (2016). A novel molecular approach for tracing terrigenous dissolved organic matter into the deep ocean. Global Biogeochemical Cycles, 30(5), 689-699. https://doi.org/10.1002/2015GB005320.
- Megonigal, J.P., Hines, M.E., and Visscher, P.T. (2004). Anaerobic metabolism: linkages to trace gases and aerobic processes. Biogeochemistry.
- Meier, H.E.M., Vali, G., Naumann, M., Eilola, K., Frauen, C. (2018). Recently accelerated oxygen consumption rates amplify deoxygenation in the Baltic Sea. Journal of Geophysical Research Oceans, 123(5), 3227-3240. https://doi.org/10.1029/2017JC013686.
- Melet, A.V., Hallberg, R., Marshall, D.P. (2021). Chapter 2: The role of ocean mixing in the climate system. In: Ocean Mixing: Drivers, Mechanisms, and Impacts, M. Meredith, A. Naveira Garabato, eds. Elsevier, pp. 5-34. https://doi.org/10.1016/B978-0-12-821512-8.00009-8.
- Meyers, M.T., Cohlan, W.P., Carpenter, E.J., Kimmerer, W.J. (2019). Effect of ocean acidification on the nutritional quality of marine phytoplankton for copepod reproduction. PLOS ONE, 14(5): e0217047. https://doi.org/10.1371/journal.pone.0217047.
- Middelburg, J.J. (2018). Reviews and syntheses: to the bottom of carbon processing at the seafloor, Biogeosciences, 15, 413-427. https://doi.org/10.5194/bg-15-413-2018.
- Middelburg, J.J., and Levin, L.A. (2009). Coastal hypoxia and sediment biogeochemistry, Biogeosciences, 6, 1273-1293. https://doi.org/10.5194/bg-6-1273-2009.
- Moore, C.M., Mills, M.M., Arrigo, K.R., and others (2013). Processes and patterns of oceanic nutrient limitation. Nature Geoscience 6, 701-710. https://doi.org/10.1038/ngeo1765.
- Mossman, H.L., Pontee, N., Born, K., Lawrence, P.J., Rae, S., Scott, J., Serato, B., Sparkes, R.B., Sullivan, M.J., and Dunk, R.M. (2012). Rapid carbon accumulation at a saltmarsh restored by managed realignment far exceeds carbon emitted in site construction. BioRxiv, pp. 2021-10. https://doi.org/10.1111/j.1365-2664.2012.02198.x.
- Nellemann, C., and others (2009). Blue Carbon: the role of healthy oceans in binding carbon. United Nations Environment Programme, GRIDArendal. https://wedocs.unep.org/20.500.11822/7772.
- Passow, U., Carlson, C.A. (2012). The biological pump in a high CO2 world. Marine Ecology Progress Series, 470, 249-271. https://doi.org/10.3354/meps09985.
- Pernet, F., Dupont, S., Gattuso, J.-P., Metian, M., and Gazeau, F. (2025). Cracking the myth: Bivalve farming is not a CO2 sink. Rev. Aquac., 17: e12954. https://doi.org/10.1111/raq.12954.
- Petillon, J., Potier, S., Carpentier, A., and Garbutt, A. (2014). Evaluating the success of managed realignment for the restoration of salt marshes: Lessons from invertebrate communities. Ecological Engineering, 69, pp. 70-75. https://doi.org/10.1016/j.ecoleng.2014.03.085.
- Phillips, B.F., Booth, J.D., Cobb, J.S., Jeffs, A.G., and McWilliam, P. (2006). Larval and Postlarval Ecology. In Lobsters: Biology, Management, Aquaculture and Fisheries (pp. 231-262). John Wiley & Sons, Ltd. https://doi.org/10.1002/9780470995969.ch7.
- Pörtner, H.-O., and others (2023). Overcoming the coupled climate and biodiversity crises and their societal impacts. Science, 380 (6642). https://doi.org/10.1126/science.abl4881.
- Poulton, A.J. (2021). Shunt or Shuttle. Nature Geoscience, 14, 181-183. https://doi.org/10.1038/s41561-021-00718-z.
- Queirós, Ana M., and others (2023). Identifying and protecting macroalgae detritus sinks toward climate change mitigation. Ecological Applications, 33.3: e2798.
- Roth, F., Broman, E., Sun, X., Bonaglia, S., Nascimento, F., Prytherch, J., Brüchert, V., Lundevall Zara, M., Brunberg, M., Geibel, M.C., and Humborg, C. (2023). Methane emissions offset atmospheric carbon dioxide uptake in coastal macroalgae, mixed vegetation and sediment ecosystems. Nature Communications, 14(1), p. 42. https://doi.org/10.1038/s41467-022-35673-9.
- Regnier, P., Resplandy, L., Najjar, R. G., and Ciais, P. (2022). The land-to-ocean loops of the global carbon cycle. Nature, 603(7901), 401-410. https://doi.org/10.1038/s41586-021-04339-9.
- Ren, X., Xu, J., Wang, H., Liu, M., Liu, X., Li, Y., and Wu, X. (2024). The depositional characteristics of turbidity current in Manila Trench: Sediment provenance, geochemical elements and organic carbon burial. Journal of Asian Earth Sciences, 264, 106074. https://doi.org/10.1016/j.jseaes.2024.106074.
- Saba, G.K., Burd, A.B., Dunne, J.P., Hernández-León, S., Martin, A.H., Rose, K.A., Salisbury, J., Steinberg, D.K., Trueman, C.N., Wilson, R. W., and Wilson, S.E. (2021). Toward a better understanding of fish-based contribution to ocean carbon flux. Limnology and Oceanography, 66(5), 1639-1664. https://doi.org/10.1002/lno.11709.
- Sabine, C. (2021). The role of the ocean in the global carbon cycle. In: Integrated Ocean Carbon Research: A summary of ocean carbon research, and vision of coordinated ocean carbon research and observations for the next decade. (Eds Wanninkof, Sabine, and Arico). IOC Technical Series 158, Paris, UNESCO, 7-9. https://doi.org/10.25607/h0gj-pq41.
- Saderne, V., Geraldi, N.R., Macreadie, P.I., and others (2019). Role of carbonate burial in Blue Carbon budgets. Nat Commun, 10, 1106. https://doi.org/10.1038/s41467-019-08842-6.
- Sanford, M.P. (2009). Valuating Mangrove Ecosystems as Coastal Protection in Post-Tsunami South Asia. Natural Areas Journal, 29(1), 91-95. https://doi.org/10.3375/043.029.0111.
- I.R. Santos, D.J. Burdige, T.C. Jennerjahn, S. Bouillon, A. Cabral, O. Serrano, T. Wernberg, K. Filbee-Dexter, J.A. Guimond, J.J. Tamborsk (2009). The renaissance of Odum's outwelling hypothesis in 'Blue Carbon' science. Estuarine, Coastal and Shelf Science, volume 255, 5 July 2021, 107361. https://doi.org/10.1016/j.ecss.2021.107361.
- Schmidtko, S., Stramma, L., Visbeck, M. (2017). Decline in global oceanic oxygen content during the past five decades. Nature, 542, 335-339. https://doi.org/10.1038/nature21399.
- Schmoker, C., Hernandez-Leon, S., Calbert, A. (2013). Microzooplankton grazing in the oceans: impacts, data variability, knowledge gaps and future directions. Journal of Plankton Research, 35(4), 691-706. https://doi.org/10.1093/plankt/fbt023.
- Shi, D., Zu, Y., Hopkinson, B.M., Morel, F.M.M. (2010). Effect of ocean acidification on iron availability to marine phytoplankton. Science, 327 (5966), 676-679. https://doi.org/10.1126/science.1183517.
- Siegel, D.A., De Vries, T., Cetinic, I., Bisson, K.M. (2023). Quantifying the ocean's biological pump and its carbon cycle impacts on global scales. Annual Review of Marine Science, 15, 329-356. https://doi.org/10.1146/annurev-marine-040722-115226.
- Suttle, C. (2007). Marine viruses - major players in the global ecosystems. Nature Reviews Microbiology, 5, 801-812. https://doi.org/10.1038/nrmicro1750.
- Tagliabue, A., Kwiatkowski, L., Bopp, L., and others (2021). Persistent uncertainties in ocean net primary production climate change projections at regional scales raise challenges for assessing impacts on ecosystem services. Front. Climate, 3. https://doi.org/10.3389/fclim.2021.738224.
- Talling, P.J., Hage, S., Baker, M.L., Bianchi, T.S., Hilton, R.G., and Maier, K.L. (2024). The Global Turbidity Current Pump and Its Implications for Organic Carbon Cycling. Annual Review of Marine Science, volume 16, 105-133. https://doi.org/10.1146/annurev-marine-032223-103626.
- Taucher, J., Haunost, M., Boxhammer, T., Bach, L.T., Alguero-Muniz, M., Riebesell, U. (2017). Influence of ocean acidification on plankton community structure during a winter-to-summer succession: An imaging approach indicates that copepods can benefit from elevated CO2 via indirect food web effects. PLOS ONE, 12(2): e0169737. https://doi.org/10.1371/journal.pone.0169737.
- Trevathan-Tackett, S.M., Callahan, D.L., Connolly, R.M., and Macreadie, P.I., (2023). Dual indicators link geochemistry to microbiota in blue carbon soils. Estuarine, Coastal and Shelf Science, 285, p. 108307.
- Tsunogai, S., Watanabe, S., and Sato, T. (1999). Is there a "continental shelf pump" for the absorption of atmospheric CO2? Tellus B: Chemical and Physical Meteorology, 51(3), 701-712. https://doi.org/10.3402/tellusb.v51i3.16468.
- Underwood, G.J., Dumbrell, A.J., McGenity, T.J., McKew, B.A., and Whitby, C. (2022). The microbiome of coastal sediments. The marine microbiome, pp. 479-534. https://doi.org/10.1007/978-3-030-90383-1_12.
- UNFCCC (2024). Nationally determined contributions under the Paris Agreement. Synthesis report by the secretariat. Conference of the Parties serving as the meeting of the Parties to the Paris Agreement Sixth session, Baku, 11 to 22 November 2024. FCCC/PA/CMA/2024/10.
- Wasserman, J., Human, L.R.D., and Adams, J.B. (2023). Blue carbon stocks in southern Africa's Endangered seagrass Zostera capensis. Estuarine, Coastal and Shelf Science, 284, p. 108296. https://doi.org/10.1016/j.ecss.2023.108296.
- Williamson, P., and Gattuso, J.-P. (2022). Carbon removal using coastal Blue Carbon ecosystems is uncertain and unreliable, with questionable climatic cost-effectiveness. Frontiers in Climate, 4. https://doi.org/10.3389/fclim.2022.853666.
- Williamson, P., R. W. Schlegel, J.-P. Gattuso, J.E. Andrews, and T.D. Jickells (2024). Climate benefits of saltmarsh restoration greatly overstated by, Mason, and others. Global Change Biology, 30: e17525. https://doi.org/10.1111/gcb.17525.
- Wilson, R.J., Speirs, D.C., Sabatino, A., and Heath, M.R. (2018) A synthetic map of the north-west European Shelf sedimentary environment for applications in marine science. Earth System Science Data, 10, 109-130. https://doi.org/10.5194/essd-10-109-2018.
- Wolf-Gladrow, D.A., Rost, B. (2014). Ocean Acidification and oceanic carbon cycling. In: Global Environmental Change: Handbook of Global Environmental Pollution, vol 1., B. Freedman, ed. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-5784-4_79.
- Zarate-Barrera, T.G., Maldonado, J.H. (2015). Valuing Blue Carbon: Carbon Sequestration Benefits Provided by the Marine Protected Areas in Colombia. PLoS ONE, 10(5): e0126627. doi:10.1371/journal. pone.0126627.