Macroalgae, not including phytoplankton
Writing team: Elizabeth J. Cottier-Cook (coordinating author), Juliet Brodie, Sophie Corrigan, Olivier De Clerck, Phaik-Eem Lim, Christine Maggs, Carmen Mifsud (lead member), Felicia Mogo (co-lead member), Flower Msuya, Franciane Pellizzari, Liu Tao, Heroen Verbruggen, Thomas Wernberg, Joanna Wilbraham and Nair Yokoya.
Key points
- Rapidly warming oceans, combined with anthropogenic stressors, will lead to further significant losses of certain macroalgal species and changes in the distribution patterns of others.
- The distribution of temperate and warmer water species is likely to shift poleward into temperate boreal and deeper waters, conditional on dispersal capacity and substrate availability.
- The incidence of invasive macroalgal blooms and turfs is likely to increase, leading to changes in ecosystem function.
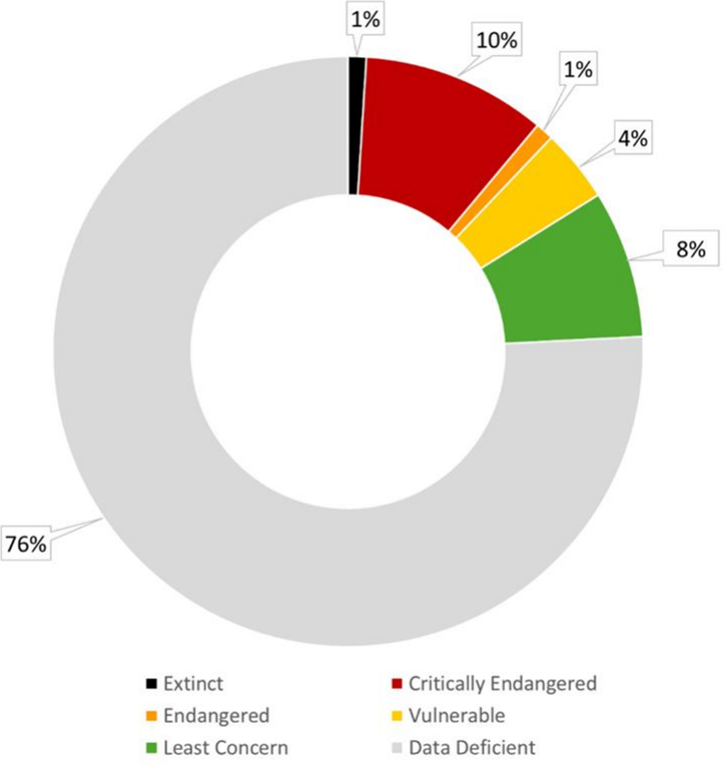
- Less than 1% of macroalgal species described to date have been evaluated for their extinction risk using the International Union for Conservation of Nature (IUCN) Red List of Threatened Species assessment, and nearly 76% of the 99 macroalgal entries on the List are Data Deficient. This significant gap poses substantial challenges for assessing risk status.
- Significant gaps in taxonomic and distribution knowledge for many macroalgae is impeding progress in the effective monitoring of changes in population size and geographical range.
1. Introduction
The present subchapter contains a presentation of the overall status of red, green and brown macroalgae (commonly known as seaweed) and changes in the period 2018-2024. Here, the macroalgae are considered in their own subchapter, having been considered as a food source and as an ecosystem in the first World Ocean Assessment and included with marine plants in the second World Ocean Assessment. Up to 2024, only 99 macroalgae, from a total of over 11,400 species described Ref 26 are included in the International Union for Conservation of Nature (IUCN) Red List Ref 34, and 76% of these are categorized as Data Deficient (figure I). Recognizing macroalgae in the third World Ocean Assessment is, therefore, timely and critically important, given that the maximum potential area of ocean that seaweeds can occupy is an estimated 6 million to 7 million km2 Ref 15. Seaweeds also play a crucial role in marine ecosystem functioning, and many species are being significantly affected by climate change and other anthropogenic stressors Ref 8.
Figure I Categorization of 99 macroalgal species on the International Union for Conservation of Nature Red List, reported from the Pacific North-West, the Pacific South-East and the Pacific South-West
2. Environmental change since the second World Ocean Assessment
Changes in overall status
Red macroalgae
Red macroalgae (Rhodophyta) occur from the poles to the tropics and from the intertidal zone to 290 m deep Ref 45. Most red macroalgae are marine (97%) Ref 67; they are habitat-forming (e.g. rhodolith beds; see figure II) and are integral components of other habitats (e.g. coral reefs and temperate and arctic rocky reefs) (see sect. 4, subchap. 5D). Red macroalgae provide ecosystem services, including by serving as breeding, spawning and nursery grounds for ecologically and commercially important marine species Ref 59. They are also harvested and/or farmed, primarily in South-East Asia for their gelling and emulsifying colloidal agents and in East Asia for food and aquaculture feed Ref 9 (see subsect. 5A, subchap. 1D). The maximum potential area for red macroalgae globally is estimated as 6.69 million km2 Ref 15, including 4.12 million km2 of rhodolith beds Ref 22.
Currently (as of 2024), 7,698 red macroalgal species have been described Ref 26, although high levels of (pseudo-) cryptic diversity mean that many still need to be described and that new species are continually being discovered Ref 44. Since the publication of the second World Ocean Assessment, two new orders have been established, Corallinapetrales Ref 35 and Inkyuleeales Ref 14. Many larger genera (e.g. Pyropia) have been split into smaller units to better reflect their evolutionary relationships Ref 76 and many species have been moved between genera on the basis of recent research Ref 30.
The role of coralline red algae in carbon sequestration remains contested, as the calcification process also releases CO2. New research suggests that coralline macroalgae recycle 40% of the carbon from calcification into photosynthesis Ref 50. Given the longevity of coralline macroalgae and the extent of rhodolith beds, globally they could represent an important store of carbon (400 to 38,000 tons of carbonate deposits) Ref 63 (see subsect. 5B, chap. 1).
Figure II Rhodolith (maerl) bed, Ireland

Green macroalgae
Green macroalgae, most of which belong to the class Ulvophyceae (Chlorophyta), consist of 1,900 species and are distributed worldwide Ref 17. They are typically found in shallow waters, although species have been recorded at depths greater than 210 m Ref 46. They provide a wide variety of ecosystem services, including an important food source for marine grazers, nutrient cycling and reef building (e.g. Halimeda spp.). Although the total coverage of green macroalgae has not been estimated, the global coverage associated with soft or sandy and coral habitats is estimated at 1.2 million km2 Ref 51. Halimeda species produce reef-like habitats (bioherms) and their carbonate production ranges from 0.8 to over 17,500 g per m2 per year in the Indo- Pacific Ref 62. These habitats are now considered to be major contributors to carbonate production in tropical and subtropical regions Ref 62.
A few opportunistic Ulvaspecies can form extensive blooms (known as "green tides"), which have increased in frequency and can cause biodiversity loss, health risks and a reduction in ecosystem services in the affected area Ref 65 Ref 36. The introduced Halimeda incrassata has also spread rapidly in shallow subtidal habitats of the western Mediterranean Sea (Vergruggen and Ballesteros, 2024).
Progress in resolving evolutionary relationships in green macroalgae is continuing. Since the publication of the second World Ocean Assessment, studies have indicated that not all green macroalgae of the class Ulvophyceae may descend from a common evolutionary ancestor Ref 12 Ref 28. Bryopsidales is likely to be more closely related to Chlorophyceae, a class of predominantly freshwater green microalgae, than to the remaining Ulvophyceae.
Several studies focused on local diversity of green macroalgae have highlighted the discovery of many undescribed taxa Ref 41 Ref 68, particularly in tropical regions. The sequencing of Ulva types has resulted in several name changes Ref 30 Ref 29 Ref 23, with U. fenestrata being the correct name of the temperate species often reported as U. lactuca, and U. lactuca being the correct name for the pan-tropical U. fasciata Ref 31.
Brown macroalgae
The brown macroalgae or Phaeophyceae (Ochrophyta) provide several ecosystem services, including food and habitat provision, nutrient cycling and carbon fixation, with an estimated value of approximately 500 billion dollars globally Ref 16. The number of brown macroalgal species has been estimated at 1,800 Ref 26, with the vast majority inhabiting the marine environment. Their distribution is global, although they are mainly temperate and polar species, except for Sargassum and species from the order Dictyotales, which are often abundant in the tropics. They occur from the intertidal zone to a depth of 30 to 40 m, although they can extend down to depths greater than 200 m (Žuljević and others, 2016). The overall estimate of the total potential area for brown macroalgae based on distribution models is 2.68 million km2, including 0.13 million km2 for the intertidal zone Ref 15, 2.5 million km2 for subtidal zone Ref 24, and a maximum of 6,000 km2 for floating Sargassum species Ref 73 Ref 79. The latter includes more than 20 million tons of floating Sargassum extending from West Africa to the Gulf of Mexico Ref 73.
Since the publication of the second World Ocean Assessment, a novel brown macroalgal genus and species, Setoutiphycus delamareoides (Ectocarpales), has been described in Japan Ref 38. Numerous other new species have also been described, including 15 new Lobophora species Ref 66 Ref 37 and two new species of Dictyota Ref 75.
Eger and others (2023) recently estimated that six major forest-forming kelps (Ecklonia, Laminaria, Lessonia, Macrocystis, Nereocystis and Saccharina) sequester 4.9 million tons of carbon from the atmosphere annually. Increased understanding of the role of brown macroalgae in the carbon cycle has also yielded new global estimates for seaweed-derived particulate carbon export below a depth of 200 m, which total 3% to 4% of the ocean carbon sink capacity and correspond to the sequestration of 4.4 to 48.5 million tons of seaweed-derived carbon annually for 100 years Ref 21.
Major threats
Changes in climate and the rapidly warming ocean (Intergovernmental Panel on Climate Change (IPCC), 2023), together with anthropogenic stressors such as overharvesting, eutrophication, ocean acidification and pollution, are predicted to lead to an unprecedented decline in many macroalgae species. Conversely, the incidence of certain macroalgal bloom-forming species and turfs is likely to increase Ref 20 Ref 2 Ref 19 (figure III). Since the publication of the second World Ocean Assessment, reports have warned, however, that ocean warming, thermohaline and circulation change, and marine heat waves, combined with other anthropogenic stressors, have resulted in the loss of more than 90% of certain kelp species in regions of Australia, the United States of America and Mexico, with significant implications for associated biodiversity and local economies Ref 11. By the year 2100, depending on the warming scenario, brown seaweeds are predicted to lose 6 to 11% of their present estimated global coverage Ref 49, and calcified red algal distribution is estimated to decline by 26 to 44% Ref 22, the latter being more susceptible to threats such as ocean acidification than the brown seaweeds Ref 7. Furthermore, due to their extremely slow growth rates (around 1 mm per year), rhodolith beds may take centuries to millennia to recover from damage and are consequently described as a non-renewable resource Ref 1.
Seaweed loss is also predicted to increase in tropical regions (e.g. tropical red algae in South-East Asia) due to the effects of climate change and the associated pest, disease and invasive species outbreaks Ref 3 (see sect. 4, chap. 6), increased incidence of typhoons, ocean acidification Ref 55 and overharvesting Ref 47. Ranges are also likely to shift further into Arctic, temperate boreal and deeper waters Ref 43 Ref 40.
Figure III Ulva species bloom in Songosongo, United Republic of Tanzania

3. Region-specific changes
Arctic Ocean
Arctic macroalgae are experiencing significant changes due to rapid warming, declining sea ice, and increased habitat availability. Many species, particularly brown and red macroalgae, are predicted to expand their distributional ranges northward from more temperate regions, with a predicted loss of existing high-latitude macroalgae occupying similar habitats (Bringloe, and others, 2022). It is also likely that the potential calcified algal coverage will decline in the region as increasing temperatures and lower pH levels reduce the ability of these algae to calcify Ref 6.
North Atlantic Ocean, Baltic Sea, Black Sea, Mediterranean Sea and North Sea
Marine heat waves in this region have led to a marked range shift or population-level change in a number of kelp species, such as Laminaria digitata, which is predicted to undergo a poleward range contraction and experience declines in growth performance Ref 42. Declines in the fucoid Fucus vesiculosus and increases in the filamentous turfing algae Pylaiella littoralis and Ectocarpus siliculosus have been attributed to increased eutrophication in the Baltic Sea Ref 27. In the Mediterranean Sea, Fucus virsoides has seen a significant decline Ref 13, and species that favour warmer waters (i.e. invasive species from tropical and subtropical regions, e.g. Rugulopteryx okamurae) have increased and spread Ref 18 Ref 74 (see sect. 4, chap. 6).
South Atlantic Ocean and wider Caribbean
Increases in macroalgal cover on hard coral reefs have been reported in the Mexican Caribbean Ref 10 and in floating, invasive, "pelagic" Sargassum species, which have extended beyond the Sargasso Sea (see subchap. 5Q) (Wang and others 2019) and proliferated in the tropical Atlantic between West Africa, Brazilian oceanic islands (Pellizzari and others 2020) and South America (Robledo and others, 2021; Sissini and others 2017) (see sect. 4, chap. 6).
Indian Ocean, Arabian Sea, Bay of Bengal, Red Sea, Gulf of Aden and Persian Gulf
As in the Caribbean, more temperature-tolerant macroalgae are becoming increasingly dominant on coral reefs following incidences of coral bleaching Ref 25.
North Pacific Ocean
Twenty red macroalgal species are reported on the IUCN Red List for this region and all are endemic to the Republic of Korea (North-West Pacific). Of these species, 4 are categorized as Least Concern and 16 are categorized as Data Deficient Ref 34. The green macroalga Codium spinulosum (Bryopsidales) and three species of brown macroalgae are also endemic to the Republic of Korea Ref 77 and are categorized as Data Deficient Ref 34.
South Pacific Ocean
The IUCN Red List contains 58 red macroalgal species found in the South Pacific Ocean. They include one Extinct species from the South-West Pacific and 57 species from the South-East Pacific, categorized as Critically Endangered (6), Vulnerable (3), Least Concern (4) and Data Deficient (44) Ref 34. The brown macroalga Nereia lophocladia is endemic to New South Wales, Australia Ref 48, in the South-West Pacific, and two green macroalgal species, Rhizoclonium robustum (Cladophorales) and Entocladia thivyae, are both endemic to the Galapagos Islands in the South-East Pacific Ref 61. All three species are categorized as Data Deficient Ref 34. Fifteen species of brown macroalgae from the South-East Pacific are thought to be endemic to the Galapagos and are categorized as Endangered (1), Critically Endangered (4), Vulnerable (1) and Data Deficient (9) in the IUCN Red List Ref 34.
Southern Ocean
Records have recently been compiled for this region, which include 151 macroalgal species (85 Rhodophyta, 34 Ochrophyta and 32 Chlorophyta) in Antarctica, in the South Shetland Islands Ref 54, King George Island and Adelaide Island Ref 53. Distributional shifts and a reduction in the endemism rate have been reported in this region Ref 58, and, according to a recent modelling study, there is a high potential for macroalgal expansion into this region as a consequence of climate change Ref 49.
4. Key remaining knowledge gaps
Despite the substantial evidence showing that macroalgal species and their habitats are declining, significant numbers of species remain undescribed, undiscovered Ref 26 and/or inadequately protected or conserved. The lack of a sound taxonomic framework for identifying and monitoring species hampers efforts to determine their distribution, threats and changes to status Ref 5, as well as efforts to implement effective conservation, protection and restoration strategies. It also presents significant challenges for assessing species for the IUCN Red List, which can raise awareness and assist in the global conservation of a species, as clearly shown by the fact that less than 1% of macroalgae known are listed from a very limited number of regions globally. The contribution of many seaweeds (e.g. rhodolith beds and kelp forests) to larger-scale processes such as carbon sequestration Ref 63 also requires further investigation (Intergovernmental Oceanographic Commission (IOC- UNESCO), 2024). There is urgent need, therefore, to (a) strengthen data-driven conservation efforts to ensure that losses in macroalgal diversity are halted and habitats restored whenever possible; (b) ensure that range shifts are reported and vulnerable species protected; (c) identify undescribed species using the latest molecular approaches; and (d) ensure that the role of macroalgae in large-scale ocean processes is more widely understood.
References
- Barbera, C., Bordehore, C., Borg, J.A., Glémarec, M., Grall, J., Hall-Spencer, J.M., De La Huz, C.H., Lanfranco, E., Lastra, M., Moore, P.G. and Mora, J. (2003). Conservation and management of northeast Atlantic and Mediterranean maerl beds. Aquatic conservation: marine and freshwater ecosystems, vol. 13, pp. S65-S76.
- Bermejo, R., Green-Gavrielidis, L. and Gao, G. (2023). Macroalgal blooms in a global change context. Frontiers in Marine Science, vol., pp. 1204117, 2023.
- Brakel, J., Sibonga, R., Dumilag, R.V., Montalescot, V., Campbell, I., Cottier-Cook, E.J., Ward, G., Le Masson, V., Liu, T., Msuya, F.E., Brodie, J., Lim, P.E. and Gachon, C.M.M. (2021). Exploring, harnessing, and conserving genetic resources towards a sustainable seaweed aquaculture. Plants, People, Planet, vol. 3, pp. 337-349.
- Bringloe, T.T., Wilkinson, D.P., Goldsmit, J., Savoie, A.M., Filbee-Dexter, K., Macgregor, K.A., Howland, K.L., McKindsey, C.W., Verbruggen, H. (2022). Arctic marine forest distribution models showcase potentially severe habitat losses for cryophilic species under climate change. Global Change Biology, vol. 28, pp. 3711-3727.
- Brodie, J., Wilbraham, J., Maggs, C.A., Baldock, L., Bunker, F., Mieszkowska, N., Scanlan, C., Tittley, I., Wilkinson, M. and Yesson, C. (2023). Red List for British seaweeds: evaluating the IUCN methodology for non-standard marine organisms. Biodiversity Conservation, vol. 32, pp. 3825-3843.
- Chan, P.T.W, Halfar, J., Adey, W.H., Lebednik, P.A., Steneck, R., Norley, C.J.D. and Holdsworth, D.W. (2020). Recent density decline in wild-collected subarctic crustose coralline algae reveals climate change signature. Geology, vol. 48, pp. 226-230.
- Cornwall, C.E., Harvey, B.P., Comeau, S., Cornwall, D.L., Hall-Spencer, J.M., Pena, V., Wada, S. and Porzio, L. (2021). Understanding coralline algal responses to ocean acidification: Meta-analysis and synthesis. Global Change Biology, vol. 28, pp. 362-374.
- Cottier-Cook, E. J., Lim, P. E., Mallinson, S., Yahya, N., Poong, S., Wilbraham, J., Nagabhatla, N., Brodie, J. (2023). Striking a balance: wild stock protection and the future of our seaweed industries. United Nations University-Insittute on Comparative Regional Integration Studies (UNU-CRIS) Policy Brief [Online].
- Cottier-Cook, E.J., Nagabhatla, N., Asri, A., Beveridge, M., Bianchi, P., Bolton, J., Bondad-Reantaso, M.G., Brodie, J., Buschmann, A., Cabarubias, J., Campbell, I., Chopin, T., Critchley, A., De Lombaerde, P., Doumeizel, V., Gachon, C.M.M., Hayashi, L., Hewitt, C.L., Huang, J., Hurtado, A.Q., Kambey, C., Kim, G.H., Le Masson, V., Lim, P .- E., Liu, T., Malin, G., Matoju, I., Montalescot, V., Msuya, F. E., Potin, P., Puspita, M., Qi, Z., Shaxson, L., Sousa Pinto, I., Stentiford, G.D., Suyo, J. and Yarish, C. (2021). Ensuring the sustainable future of the rapidly expanding global seaweed aquaculture industry - a vision. UNU-CRIS Policy Brief [Online]
- Contreras-Silva, A.I., Tilstra, A., Migani, V., Thiel, A., Pérez-Cervantes, E., Estrada-Saldívar, N., Elias- Ilosvay, X., Mott, C., Alvarez-Filip, L., and Wild, C. (2020). A meta-analysis to assess long-term spatiotemporal changes of benthic coral and macroalgae cover in the Mexican Caribbean. Scientific Reports, vol. 10, pp. 8897.
- Dalmau, N.A., Cebrian, E., Dunga, L.V., Horta, P., Layton, C., Macaya, E.C., Mangialajo, L., Moore, P.J., Mora-Soto, A., Reddy, M.M., Smit, A.J., Spector, P., Nguyen, V.T., Verdura, J., and Pocklington, J.B. (2024). Introducing the Seaweed Specialist Group of the IUCN Species Survival Commission, Cambridge, PRESS, C. U ..
- Del Cortona, A., Jackson, C.J., Bucchini, F., Van Bel, M., D'hondt, S., Skaloud, P., Delwiche, C.F., Knoll, A.H., Raven, J.A., Verbruggen, H., Vandepoele, K., De Clerck, O. and Leliaert, F. (2020). Neoproterozoic origin and multiple transitions to macroscopic growth in green seaweeds. Proceedings of the National Academy of Sciences of the United States of America, vol. 117, pp. 2551.
- Descourvières, E., Bandelj, V., Sfriso, A., Orlando-Bonaca, M., Mačić, V., Iveša, L., Kipson, S., Gljušćić, E., Battelli, C., Moro, I., Solidoro, C., and Falace, A. (2024). Toward the first documented extinction of a marine macroalga in the Mediterranean Sea? Regional Environmental Change, vol. 24.
- Díaz-Tapia, P., Pasella, M.M., Verbruggen, H., and Maggs, C.A. (2019). Morphological evolution and classification of the red algal order Ceramiales inferred using plastid phylogenomics. Molecular Phylogenetic Evolution, vol. 137, pp. 76-85.
- Duarte, C.M., Gattuso, J .- P., Hancke, K., Gundersen, H., Filbee-Dexter, K., Pedersen, M.F., Middleburg, J.J., Burrows, M.T., Krumhansl, K.A., Wernberg, T., Moore, P., Pessarrondona, A., Orberg, S.B., Pinto, I.S., Assis, J., Queiros, A.M., Smale, D., Bekkby, T., Serrao, E.A. and Krause-Jensen, D. (2022). Global estimates of the extent and production of macroalgal forests. Global Ecology and Biogeography, vol. 31, pp. 1422-1439.
- Eger, A.M., Marzinelli, E.M., Beas-Luna, R., Blain, C.O., Blamey, L.K., Byrnes, J.E.K., Carnell, P.E., Choi, C.G., Hessing-Lewis, M., Kim, K.Y., Kumagai, N.H., Lorda, J., Moore, P., Nakamura, Y., Perez- Matus, A., Ponjtier, O., Smale, D., Steinberg, P.E., and Verges, A. (2023). The value of ecosystem services in global marine kelp forests. Nature Communications, vol. 14, pp. 1894.
- El-Manaway, I.M. and Rashedy, S.H. (2022). The ecology and physiology of seaweeds: An Overview. In: RAO, A. R. and RAVISHANKAR, G. A., eds., Sustainable Global Resources of Seaweeds. Cham, Switzerland: Springer.
- Falace, A., Alongi, G., Orlando-Bonaca, M. and Becvilacqua, S. (2024). Species loss and decline in taxonomic diversity of macroalgae in the Gulf of Trieste (Northern Adriatic sea) over the last six decades. Marine Environmental Research, vol. 202, pp. 106828s.
- Feng, Y., Xiong, Y., Hall-Spencer, J.M., Liu, K., Beardall, J., Gao, K., Ge, J., Xu, J., and Gao, G. (2023). Shift in algal blooms from micro- to macroalgae around China with increasing eutrophication and climate change. Global Change Biology, vol. 30, pp. e17018.
- Filbee-Dexter, K. and Wernberg, T. (2018). Rise of Turfs: A New Battlefront for Globally Declining Kelp Forests. Bioscience, vol. 68, pp. 64-76.
- Filbee-Dexter, K., Pessarrodona, A., Pedersen, M.F., Wernberg, T., Duarte, C.M., Assis, J., Bekkby, T., Burrows, M.T., Carlson, D.F., Gattuso, J.P., Gundersen, H., Hancke, K., Krumhansl, K.A., Kuwae, T., Middelburg, J.J., Moore, P.J., Queirós, A.M., Smale, D., Sousa Pinto, I., Suzuki, N., and Krause-Jensen, D. (2024). Carbon export from seaweed forests to deep ocean sinks. Nature Geoscience, vol. 17, pp. 552- 559.
- Fragkopoulou, E., Serrão, E.A., Horta, P.A., Koerich, G., and Assis, J. (2021). Bottom trawling threatens future climate refugia of rhodoliths globally. Frontiers in Marine Science, vol. 7, pp. 594537.
- Gabrielson, P.W., Smith, A.C., Bruno, J.F., Vision, T.J., and Brandt M. (2024). Taxonomic assessment of blade-forming Ulva species (Ulvales, Chlorophyta) in the Galápagos Archipelago, Ecuador using DNA sequencing. Botanica Marina, vol. 67, pp. 153-164.
- Gattuso, J .- P., Gentili, B., Duarte, C.M., Kleypas, J.A., Middelburg, J.J., and Antoine, D. (2006). Light availability in the coastal ocean: Impact on the distribution of benthic photosynthetic organisms and contribution to primary production. Biogeosciences Discussions, vol. 3, pp. 895-959.
- Gonzalez, K., Daraghmeh, N., Lozano-Cortés, D., Benzoni, F., Berumen, M.L. and Carvalho, S. (2024). Differential spatio-temporal responses of Red Sea coral reef benthic communities to a mass bleaching event. Scientific reports, vol. 14, pp. 24229.
- Guiry, M.D. (2024). How many species of algae are there? A reprise. Four kingdoms, 14 phyla, 63 classes and still growing. Journal of Phycology, vol. 60, pp. 214-228.
- Hall, J.R., Herkül, K., Baltar, F., Hepburn, C.D. and Martin, G. (2024). Shifts in macroalgae composition alters carbon flow in Coastal Baltic Sea ecosystems: implications for dissolved organic carbon bioavailability and flux. Frontiers in Marine Science, vol. 11, pp.
- Hou, Z., Ma, X., Shi, X., Li, X., Yang, L., Xiao, S., De Clerck, O., Leliaert, F. and Zhong, B. (2022). Phylotranscriptomic insights into a Mesoproterozoic-Neoproterozoic origin and early radiation of green seaweeds (Ulvophyceae). Nature Communications, vol. 13, pp. 1610.
- Hughey, J.R., Gabrielson, P.W., Maggs, C.A. and Mineur, F. (2021). Genomic analysis of the lectotype specimens of European Ulva rigida and Ulva lacinulata (Ulvaceae, Chlorophyta) reveals the ongoing misapplication of names. European Journal of Phycology, vol. 57, pp. 143-153.
- Hughey, J.R., Gabrielson, P.W., Maggs, C.A., Mineur, F. and Miller, K.A. (2020). Taxonomic revisions based on genetic analysis of type specimens of Ulva conglobata, U. laetevirens, U. pertusa and U. spathulata (Ulvales, Chlorophyta). Phycological Research, vol. 69, pp. 148-153.
- Hughey, J.R., Maggs, C.A., Mineur, F., Jarvis, C., Miller, K.A., Shabaka, S.H. and Gabrielson, P. (2019). Genetic analysis of the Linnaean Ulva lactuca (Ulvales, Chlorophyta) holotype and related type specimens reveals name misapplications, unexpected origins, and new synonymies (Letter). Journal of Phycology vol. 55, pp. 503-508.
- IOC-UNESCO (2024). State of the Ocean Report. Paris.
- IPCC (2023). Climate Change 2023: Synthesis Report. Geneva, Switzerland.
- IUCN (2024). The IUCN Red List of Threatened Species [Online]. Available at https://www.iucnredlist.org/en.
- Jeong, S.Y., Nelson, W.A., Sutherland, J.E., Peña, V., Le Gall, L., Diaz-Pulido, G., Won, B.Y., and Cho, T.O. (2021). Corallinapetrales and Corallinapetraceae: a new order and family of coralline red algae including Corallinapetra gabrielii comb. nov. Journal of Phycology, vol. 57, pp. [1-14] 849-862R.
- Joniver, C.F.H., Photiades, A., Moore, P.J., Winters, A.L., Woolmer, A., and Adams, J.M.M. (2021). The global problem of nuisance macroalgal blooms and pathways to its use in the circular economy. Algal Research, vol. 58, pp. 102407.
- Jurejan, N., Draisma, S., Vieira, C., Sun, Z., Avakul, P., Klomjit, A., Chirapart, A. and Muangmai, N. (2024). Three new Lobophora species (Dictyotales, Phaeophyceae) from Phuket on the west coast of Thailand. Botanica Marina, vol. 67.
- Kawai, H. and Hanyuda, T. (2021). Discovery of a novel brown algal genus and species Setoutiphycus delamareoides (Phaeophyceae, Ectocarpales) from the Seto Inland Sea, Japan. Scientific reports, vol. 11, pp. 13901.
- Krause-Jensen, D., Archambault, P., Assis, J., Bartsch, I., Bischof, K., Filbee-Dexter, K., Dunton, K.H., Maximova, O., Ragnarsdottir, S.B., Sejr, M.K., Simakova, U., Spiridonov, V.A., Wegeberg, S., Winding, M.H.S., and Duarte, C.M. (2020). Imprint of Climate Change on Pan-Arctic Marine Vegetation. Frontiers in Marine Science, vol. 7.
- La Guardia, L.C., Filbee-Dexter, K., Reimer, J., Macgregor, K.A., Garrido, I., Singh, R.K., Bélanger, S., Konar, B., Iken, K., Johnson, L.E., Archambault, P., Sejr, M.K., Søreide, J.E., and Mundy, C.J. (2023). Increasing depth distribution of Arctic kelp with increasing number of open water days with light. Elementa: Science of the Anthropocene, vol. 11, pp. 00051.
- Lagourgue, L., Gobin, S., Maele, B., Sylvette, V., Bonneville, C., Thierry, J., Simon, V.W. and Payri, C.E. (2022). Ten new species of Ulva (Ulvophyceae, Chlorophyta) discovered in New Caledonia: genetic and morphological diversity, and bloom potential. European Journal of Phycology, vol. 57, pp. 458-478.
- Leathers, T., King, N.G., Foggo, A., Smale, D.A. (2024). Marine heatwave duration and intensity interact to reduce physiological tipping points of kelp species with contrasting thermal affinities. Annals of Botany, vol. 133, pp. 51-59.
- Lebrun, A., Comeau, S., Gazeau, F., and Gattuso, J.P. (2022). Impact of climate change on Arctic macroalgal communities. Global and Planetary Change, vol. 219, pp. 103980.
- Leliaert, F., Payo, D.A., Gurgel, C.F.D., Schils, T., Draisma, S.G., Saunders, G.W., Kamiya, M., Sherwood, A.R., Lin, S.M., Huisman, J.M., Le Gall, F., Anderson, R.J., Bolton, J.J., Mattio, L., Zubia, M., Spokes, T., Vieira, C., Payri Claude, E., Coppejans, E., D'Hondt, S., Verbruggen, H., and De Clerck, O. (2018). Patterns and drivers of species diversity in the Indo-Pacific red seaweed Portieria. Journal of Biogeography, vol. 45, pp. 2299.
- Littler, M.M., Littler, D.S., and Hanisak, M.D. (1991). Deep-water rhodolith distribution, productivity, and growth history at sites of formation and subsequent degradation. Journal of experimental marine biology and ecology, vol. 150, pp. 163-d182.
- Littler, M.M., Littler, D.S., Blair, S.M., and Norris, J.N. (1985). Deepest known plant life discovered on an uncharted seamount. Science, vol. 227, pp. 57-59.
- Mac Monagail, M., Cornish, L., Morrison, L., Araújo, R., and Critchley, A.T. (2017). Sustainable harvesting of wild seaweed resources. European Journal of Phycology, vol. 52, pp. 371-390.
- Mamo, L.T., Wood, G., Wheeler, D., Kelaher, B.P. and Coleman, M.A. (2021). Conservation genomics of a critically endangered brown seaweed. Journal Phycology, vol. 57, pp. 1345-1355.
- Manca, F., Benedetti-Cecchi, L., Bradshaw, C.J.A., Cabeza, M., Gustafsson, C., Norkko, A.M., Roslin, T.V., Thomas, D.N., White, L., and Strona, G. (2024). Projected loss of brown macroalgae and seagrasses with global environmental change. Nature Communications, vol. 15, pp. 5344.
- Mao, J., Burdett, H.L., and Kamenos, N.A. (2024). Efficient carbon recycling between calcification and photosynthesis in red coralline algae. Biology Letters, vol. 20, pp. 20230598.
- McNeil, M.A., Webster, J.M., Beaman, R.J., and Graham, T.L. (2016). New constraints on the spatial distribution and morphology of the Halimeda bioherms of the Great Barrier Reef, Australia. Coral Reefs, vol. 35, pp. 1343-1355.
- Murúa, P., Muñoz, L., Bustamante, D., Gauna, C., Hayashi, L., Robledo, D., Strittmatter, M., Arce, P., Westermeier, R., Müller, D.G., and Gachon, C.M.M. (2024). The first phycopathological atlas in Latin America unveils the underdocumentation of algal pathogens. Algal Research, vol. 82, pp. 103604.
- Nelson, W.A., Neill, K.F., D'archino, R., and Sutherland, J.E. (2022). Marine macroalgae of the Balleny Islands and Ross Sea. Antarctic Science, vol. 34, pp. 298-312.
- Oliveira, M.C., Pellizzari, F., Medeiros, A.S., and Yokoya, N.S. (2020). Diversity of Antarctic seaweeds. In I. Gómez, and P. Huovinen, eds., Antarctic seaweeds, 1st edition. Cham: Springer International Publishing, 23-42.
- Page, H.N., Bahr, K.D., Cyronak, T., Jewett, E.B., Johnson, M.D., and Mccoy, S.J. (2022). Responses of benthic calcifying algae to ocean acidification differ between laboratory and field settings. ICES Journal of Marine Science, vol. 79, pp. 1-11.
- Pellizzari, F., Mello, J.P.S., Osaki, V.S., Santos-Silva, M.C., Brandini, F.P., Convey, P., and Rosa, L.H. (2023). New records and updated distributional patterns of macroalgae from the South Shetland Islands and northern Weddell Sea, Antarctica. Antarctic Science, vol. 35, pp. 243-255.
- Pellizzari, F., Osaki, V.S., and Santos-Silva, M.C. (2020). New records of seaweeds and filamentous cyanobacteria from Trindade Island: an updated checklist to support conservation guidelines and monitoring of environmental changes in the southern Atlantic archipelagos. Scientia Marina, vol. 84, pp. 227-242.
- Pellizzari, F., Santos-Silva, M.C, Medeiros, A., Oliveira, M.C., Yokoya, N.S., Pupo, D. and Rosa, L. (2017). Diversity and spatial distribution of seaweeds in the South Shetland Islands, Antarctica: an updated database for environmental monitoring under climate change scenarios. Polar Biology, vol. 40, pp. 1671-1685.
- Rendina, F., Buonocore, E., Di Montanara, A.C., and Russo, G.F. (2022). The scientific research on rhodolith beds: A review through bibliometric network analysis. Ecological Informatics, vol. 70, pp. 101738.
- Robledo, D., Vazquez-Delfin, E., Freile-Pelegrin, Y., Vasquez-Elizondo, R. M., Qui-Minet, Z. N., and Salazar-Garibay, A. (2021). Challenges and Opportunities in Relation to Sargassum Events Along the Caribbean Sea. Frontiers in Marine Science, vol. 8, pp. 699664.
- Ruiz, D., and Ziemmeck, F. (2013). CDF Checklist of Galapagos Heterokonts: Brown algae and related groups - FCD Lista de especies de Heterokonta: Algas pardas y grupos relacionados de Galápagos. In Charles Darwin Foundation Galapagos Species Checklist - Lista de Especies de Galápagos de la Fundación Charles Darwin, Bungartz, F., Herrera, H., Jaramillo, P., Tirado, N., Jimenez-Uzcategui, G., Ruiz, D., Guezou, A. and Ziemmeck, F., eds. Puerto Ayora: Charles Darwin Foundation / Fundación Charles Darwin.
- Schubert, N., Alvarez-Filip, L. and Hofmann, L.C. (2023). Systematic review and meta-analysis of ocean acidification effects in Halimeda: Implications for algal carbonate production. Climate Change Ecology, vol. 4, pp. 100059.
- Schubert, N., Tuya, F., Peña, V., Horta, P.A., Salazar, V.W., Neves, P., Ribeiro, C., Otero-Ferrer, F., Espino, F., Schoenrock, K., Ragazzola, F., Olive, I., Giaccone, T., Nannini, M., Mangano, M.C., Sara, G., Mancuso, F.P., Tantillo, M.F., Bosch-Belmer, M., Martin, S., Le Gall, L., Santos, R. and Silva, J. (2024). Pink power - the importance of coralline algal beds in the oceanic carbon cycle. Nature Communications, vol. 15, pp. 8282.
- Sissini, M.N., Barreto, M.B.B., Szechy, M.T.M., De Lucena, M.B., Oliverra, M.C., Gower, J., Liu, G., De Oliveria Bastos, E., Milstein, D., Gusmao, F., Martinelli-Filho, J.E., Alves-Lima, C., Colepicolo, P., Ameka, G., De Graft-Johnson, K., Gouvea, L., Torrano-Silva, G., Nauer, F., De Castro Nunes, J.M., Barufi, J.B., Rorig, L., Riosmena-Rodriguez, R., Mello, T.J., Lotufo, L.V.C. and Horta, P.A. (2017). The floating Sargassum (Phaeophyceae) of the South Atlantic Ocean - likely scenarios. Phycologia, vol. 56, pp. 321-328.
- Smetacek, V. and Zingone, A. (2013). Green and golden seaweed tides on the rise. Nature, vol. 504, pp. 84-88.
- Sun, Z., Dao, M.D., Tran, Q.T., and Dam, D.T. (2021). A new species Lobophora tsengii sp. nov. (Dictyotales; Phaeophyceae) from Bach Long Vy (Bailongwei) Island, Vietnam. Journal of Oceanography and Limnology, vol. 39, pp. 2363-2369, 2021.
- Sutherland, J.E., Lindstrom, S., Nelson, W., Brodie, J., Lynch, M., Hwang, M.S., Choi, H .- G., Miyata, M., Kikuchi, N., Oliveira, M., Farr, T., Neefus, C., Mortensen, A., Milstein, D., and Müller, K. (2011). A new look at an ancient order: generic revision of the Bangiales. Journal of Phycology, vol. 47, pp. 1131- 1151.
- Tran, L .- A.T., Vieira, C., Steinhagen, S., Maggs, C.A., Hiraoka, M., Shimada, S., Van Nguyen, T., De Clerck, O., and Leliaert, F. (2022). An appraisal of Ulva (Ulvophyceae, Chlorophyta) taxonomy. Journal Applied Phycology, vol. 34, pp. 2689-2703.
- United Nations (2021). The second World Ocean Assessment - Volume 1, New York.
- UNEP (2016). The First Global Integrated Marine Assessment: World Ocean Assessment I.
- Verbruggen, H., and Ballesteros, E. (2024). An Atlantic origin for the introduced species Halimeda incrassata (Bryopsidales, Chlorophyta). Mediterranean Marine Science, 25(1), 55-57.
- Vieira, C., Anderson, R.J., Engelen, A.H., D'hondt, S., Leliaert, F., Payri, C., Kawai, H., and De Clerck, O. Lobophora (Dictyotales, Phaeophyceae) from the western Indian Ocean: diversity and biogeography. South African Journal of Botany, vol. 142, pp. 230-246.
- Wang, M., Hu, C., Barnes, B.B., Mitchum, G., Lapointe, B., and Montoya, J.P. (2019). The great Atlantic Sargassum belt. Science, vol. 365, pp. 83-87.
- Wesselmann, M., Hendricks, I.E., Johnson, M.D., Jorda, G., Mineur, F., and Marba, N. (2024). Increasing spread rates of tropical non-native macrophytes in the Mediterranean Sea. Global Change Biology, vol. 30, pp. e17249.
- Win, N.N.W., Hanyuda, T., Assuyuti, Y., Putri, L., Geraldino, P.J., and Kawai, H. (2024). Two new species of Dictyota (Dictyotales, Phaeophyceae), D. dimorphosa sp. nov . and D. recumbens sp. nov., based on morphological and molecular investigations. Phycologia, vol. 63, pp. 290-302.
- Yang, L .- E., Deng, Y.Y., Xu, G .- P., Russell, S., Lu, Q .- Q., and Brodie, J. (2020). Redefining Pyropia (Bangiales, Rhodophyta): four new genera, resurrection of Porphyrella and description of Calidia pseudolobata sp. nov. from China. Journal of Phycology, vol. 56, pp. 862-879.
- Yim, D. and Won, S .- Y. (2023). Codium spinulosum. The IUCN Red List of Threatened Species 2023: e.T198113266A200405856 [Online]. IUCN. Available at
- https://www.iucnredlist.org/species/198113266/200405856.
- Zhang, J., Ding, X., Zhuang, M., Wang, S., Chen, L., Shen, H., and He, P. (2019). An increase in new Sargassum (Phaeophyceae) blooms along the coast of the East China Sea and Yellow Sea. Phycologia, vol. 58, pp. 374-381.
- Žuljević, A., Peters, A.F., Nikolić, V., Antolić, B., Despalatović, M., Cvitković, I., Isajlović, I., Mihanović, H., Matijević, S., Shewring, D.M., and Canese, S. (2016). The Mediterranean deep-water kelp Laminaria rodriguezii is an endangered species in the Adriatic Sea. Marine biology, vol. 163, pp. 1-12.